

GABAergic synchronization in the limbic system and its role in the generation of epileptiform activity
- PMID: 21802488
- PMCID: PMC4878907
- DOI: 10.1016/j.pneurobio.2011.07.003
GABAergic synchronization in the limbic system and its role in the generation of epileptiform activity
Abstract
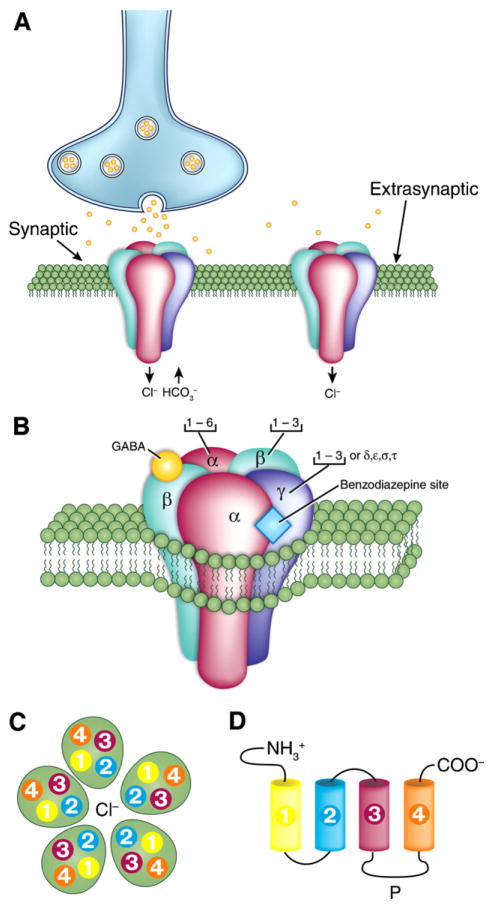
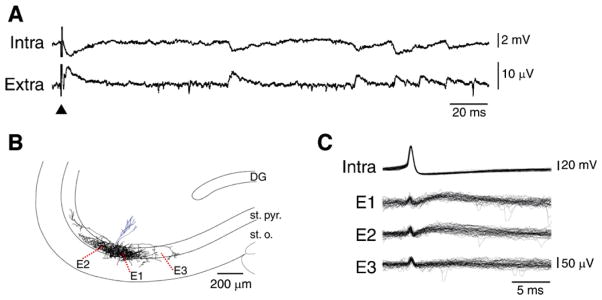
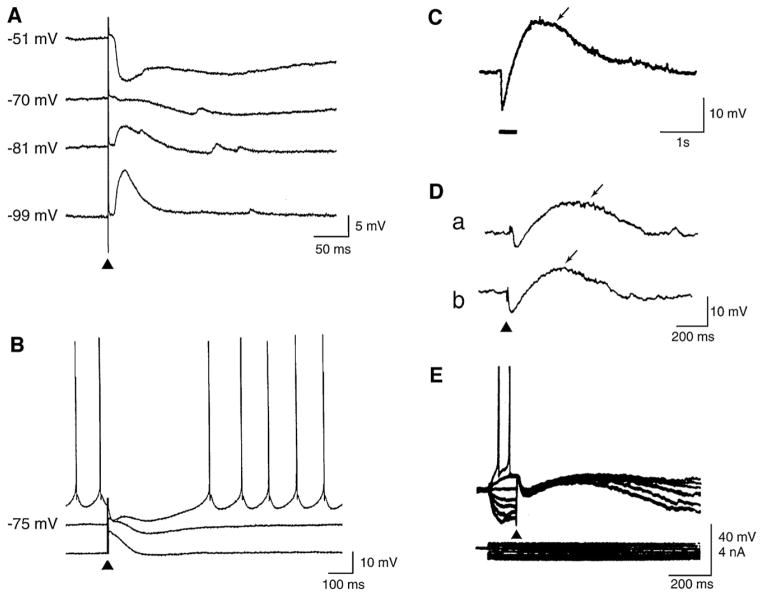
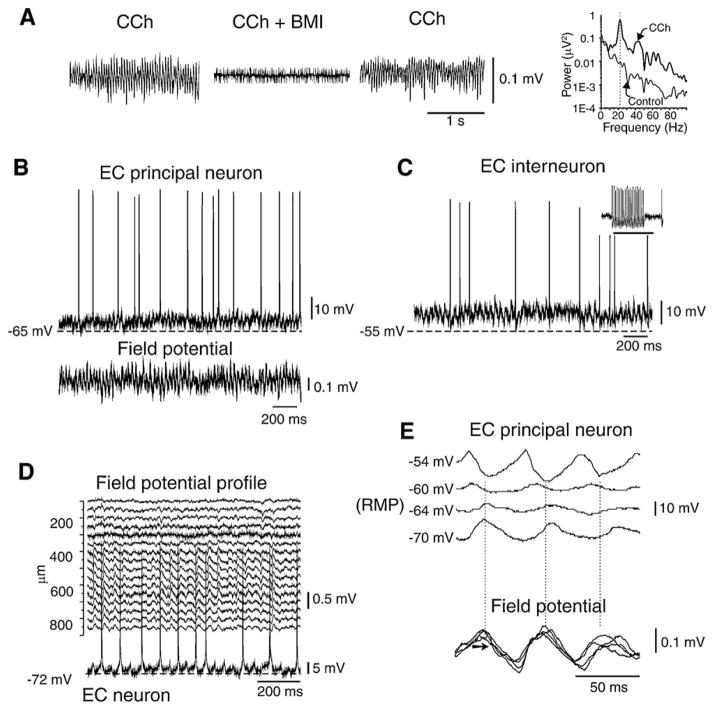
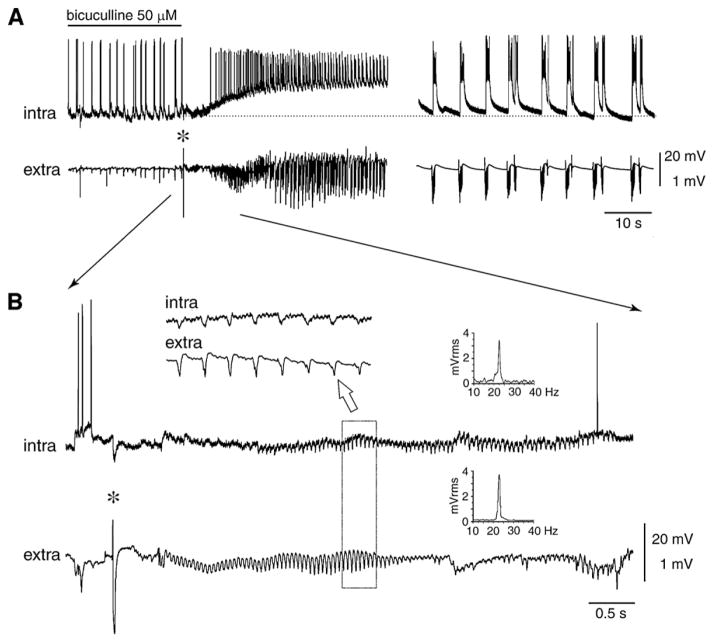
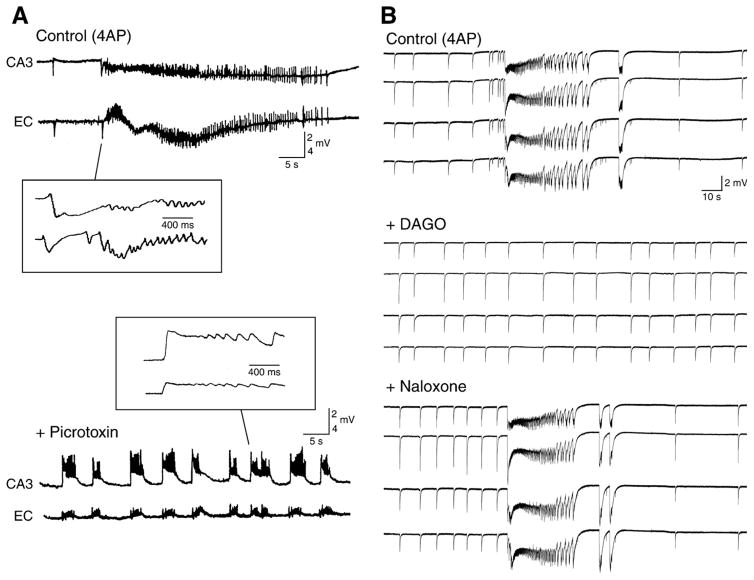
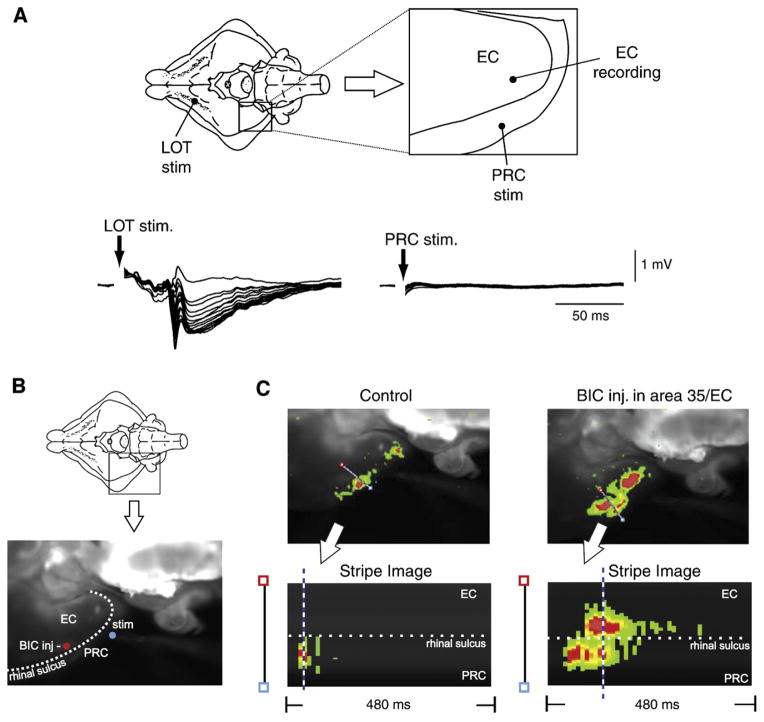
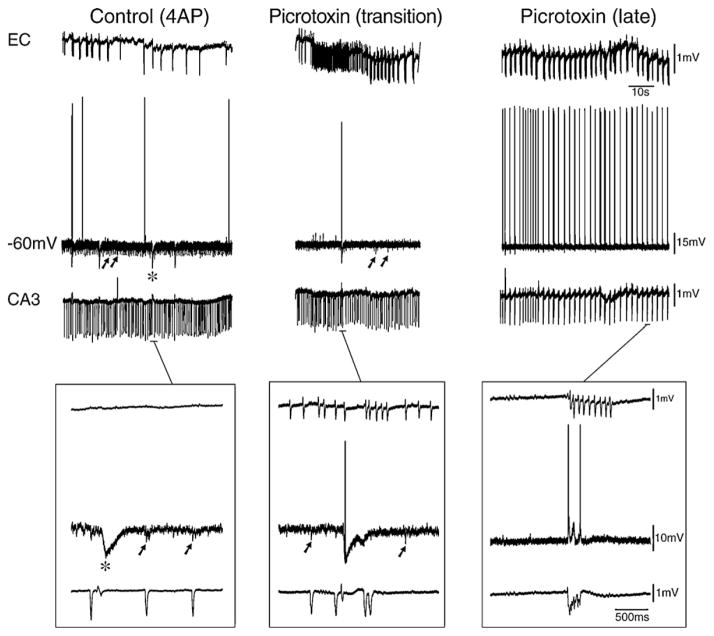
GABA is the main inhibitory neurotransmitter in the adult forebrain, where it activates ionotropic type A and metabotropic type B receptors. Early studies have shown that GABA(A) receptor-mediated inhibition controls neuronal excitability and thus the occurrence of seizures. However, more complex, and at times unexpected, mechanisms of GABAergic signaling have been identified during epileptiform discharges over the last few years. Here, we will review experimental data that point at the paradoxical role played by GABA(A) receptor-mediated mechanisms in synchronizing neuronal networks, and in particular those of limbic structures such as the hippocampus, the entorhinal and perirhinal cortices, or the amygdala. After having summarized the fundamental characteristics of GABA(A) receptor-mediated mechanisms, we will analyze their role in the generation of network oscillations and their contribution to epileptiform synchronization. Whether and how GABA(A) receptors influence the interaction between limbic networks leading to ictogenesis will be also reviewed. Finally, we will consider the role of altered inhibition in the human epileptic brain along with the ability of GABA(A) receptor-mediated conductances to generate synchronous depolarizing events that may lead to ictogenesis in human epileptic disorders as well.
Copyright © 2011. Published by Elsevier Ltd.
Figures
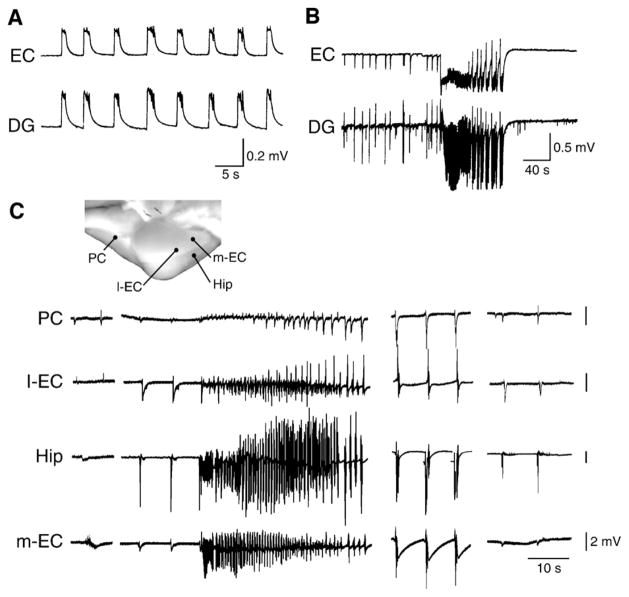
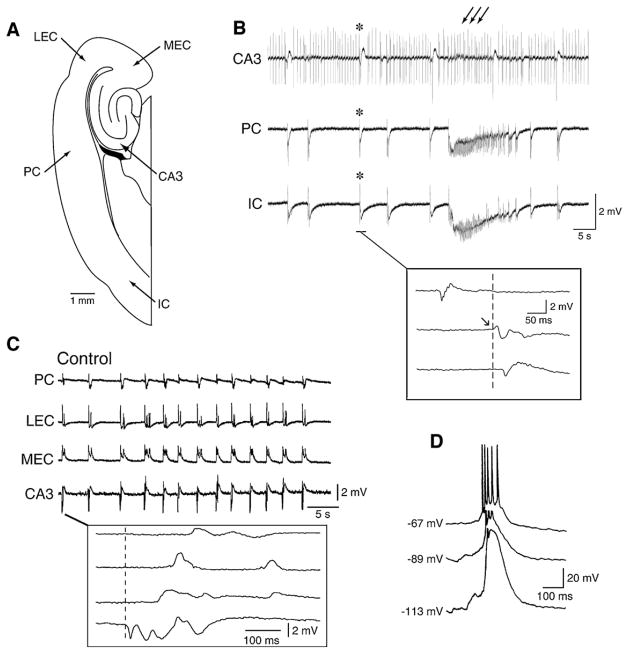
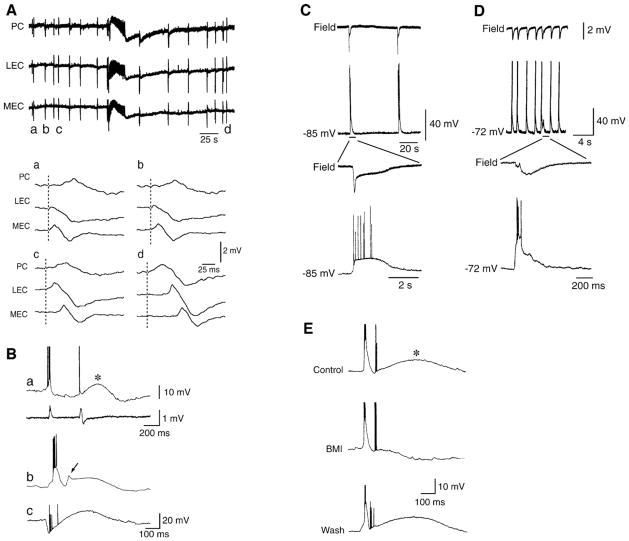
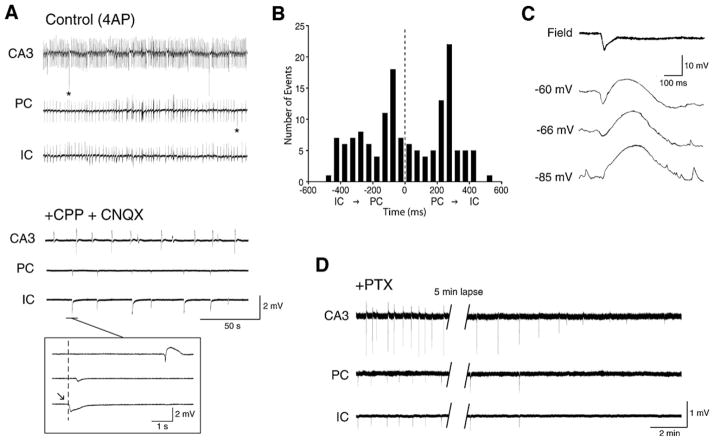
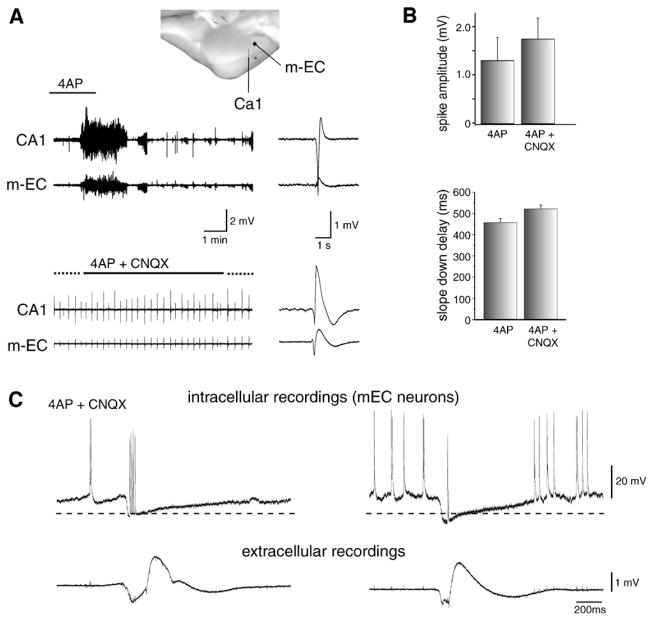
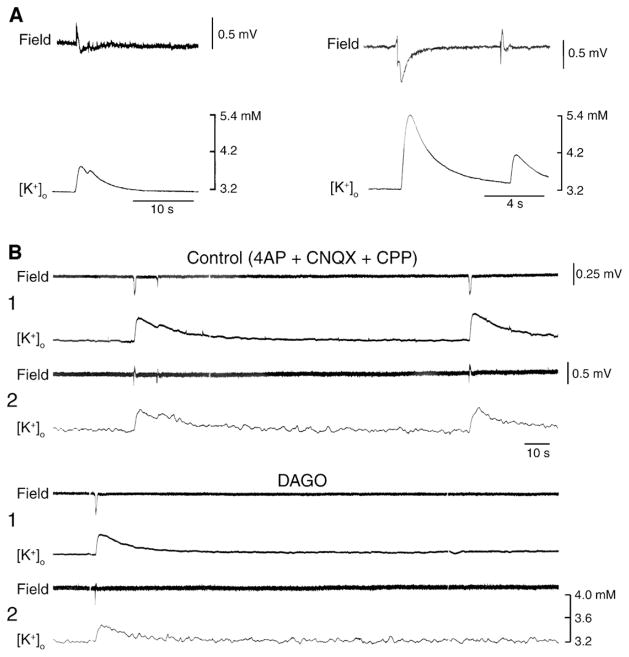
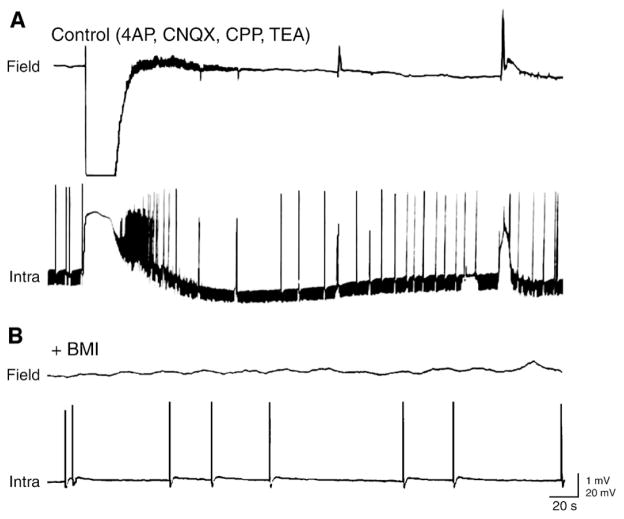
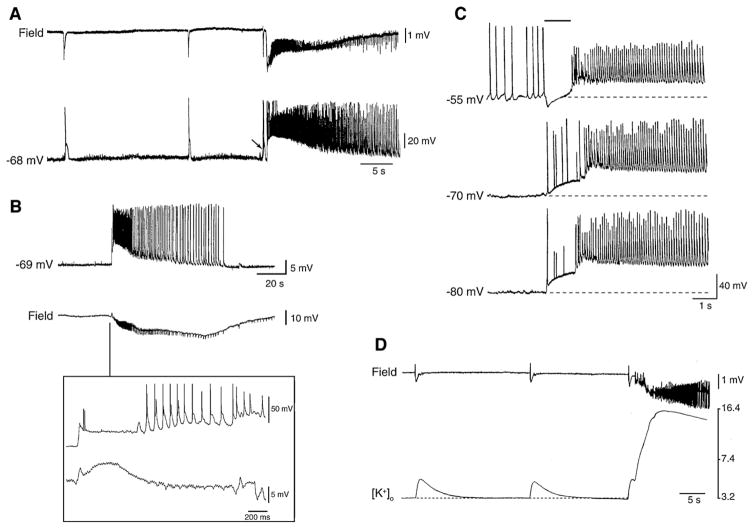
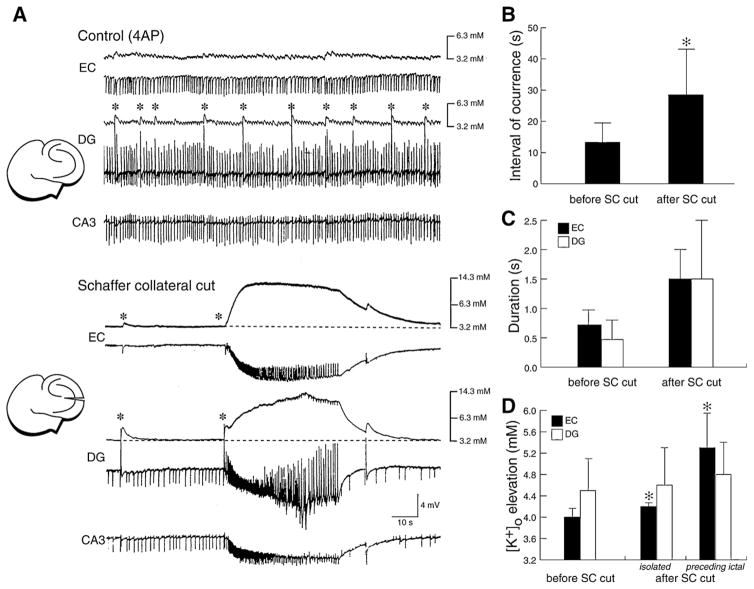
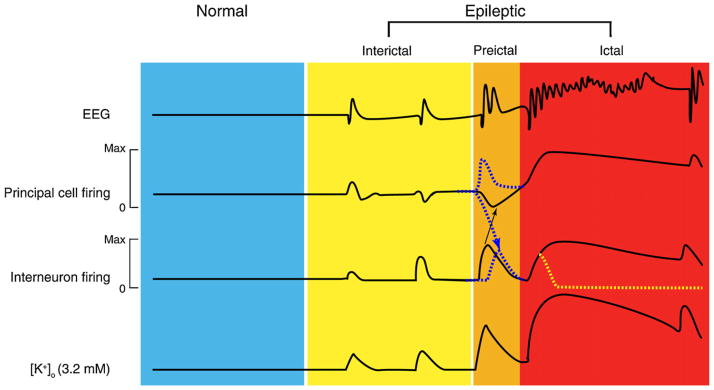
References
-
- Aicardi J. Epilepsy in children. In: Aicardi J, editor. International Review of Child Neurology Series. Raven Press; New York: 1986. p. 413.
-
- Alakuijala A, Alakuijala J, Pasternack M. Evidence for a functional role of GABA-C receptors in the rat mature hippocampus. Eur J Neurosci. 2006;23:514–520. - PubMed
-
- Alonso-Nanclares L, Garbelli R, Sola RG, Pastor J, Tassi L, Sreafico R, De Felipe J. Microanatomy of the dysplastic neocortex from epileptic patients. Brain. 2005;128:158–173. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
