

Compatibility and incompatibility in S-RNase-based systems
- PMID: 21803740
- PMCID: PMC3170157
- DOI: 10.1093/aob/mcr179
Compatibility and incompatibility in S-RNase-based systems
Abstract
Background: S-RNase-based self-incompatibility (SI) occurs in the Solanaceae, Rosaceae and Plantaginaceae. In all three families, compatibility is controlled by a polymorphic S-locus encoding at least two genes. S-RNases determine the specificity of pollen rejection in the pistil, and S-locus F-box proteins fulfill this function in pollen. S-RNases are thought to function as S-specific cytotoxins as well as recognition proteins. Thus, incompatibility results from the cytotoxic activity of S-RNase, while compatible pollen tubes evade S-RNase cytotoxicity.
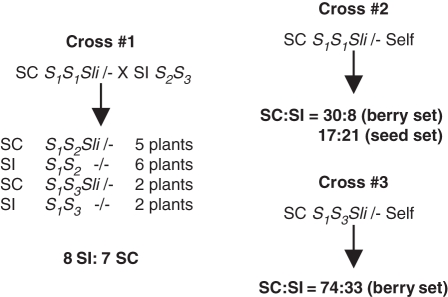
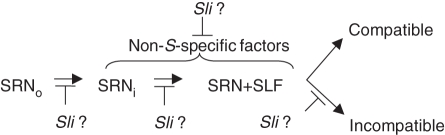
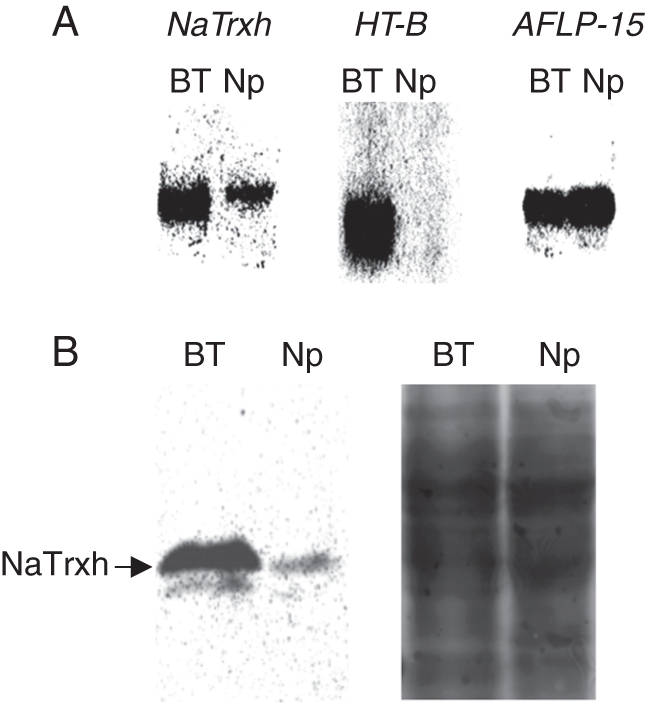
Scope: The S-specificity determinants are known, but many questions remain. In this review, the genetics of SI are introduced and the characteristics of S-RNases and pollen F-box proteins are briefly described. A variety of modifier genes also required for SI are also reviewed. Mutations affecting compatibility in pollen are especially important for defining models of compatibility and incompatibility. In Solanaceae, pollen-side mutations causing breakdown in SI have been attributed to the heteroallelic pollen effect, but a mutation in Solanum chacoense may be an exception. This has been interpreted to mean that pollen incompatibility is the default condition unless the S-locus F-box protein confers resistance to S-RNase. In Prunus, however, S-locus F-box protein gene mutations clearly cause compatibility.
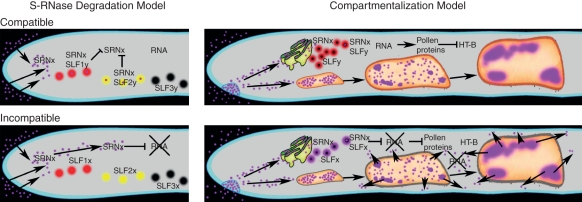
Conclusions: Two alternative mechanisms have been proposed to explain compatibility and incompatibility: compatibility is explained either as a result of either degradation of non-self S-RNase or by its compartmentalization so that it does not have access to the pollen tube cytoplasm. These models are not necessarily mutually exclusive, but each makes different predictions about whether pollen compatibility or incompatibility is the default. As more factors required for SI are identified and characterized, it will be possible to determine the role each process plays in S-RNase-based SI.
Figures
References
-
- Anderson MA, Cornish EC, Mau S-L, et al. Cloning of cDNA for a stylar glycoprotein associated with expression of self-incompatibility in Nicotiana alata. Nature. 1986;321:38–44.
-
- Ben-Naim O, Eshed R, Parnis A, et al. The CCAAT binding factor can mediate interactions between CONSTANS-like proteins and DNA. The Plant Journal. 2007;46:462–476. - PubMed
-
- Boskovic RI, Tobutt KR. Correlation of stylar ribonuclease zymograms with incompatibility alleles in sour cherry. Euphytica. 1996;90:245–250.
-
- Bredemeijer GMM, Blaas J. S-Specific proteins in styles of self-incompatible Nicotiana alata. Theoretical and Applied Genetics. 1981;59:185–190. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
