

Acetylation of heat shock protein 20 (Hsp20) regulates human myometrial activity
- PMID: 21803775
- PMCID: PMC3190820
- DOI: 10.1074/jbc.M111.278549
Acetylation of heat shock protein 20 (Hsp20) regulates human myometrial activity
Abstract
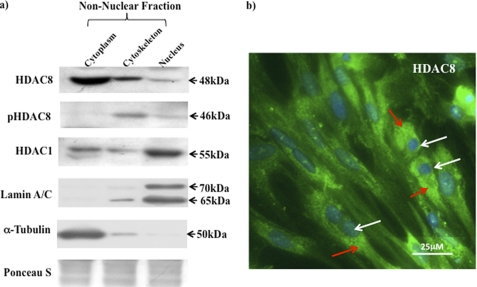
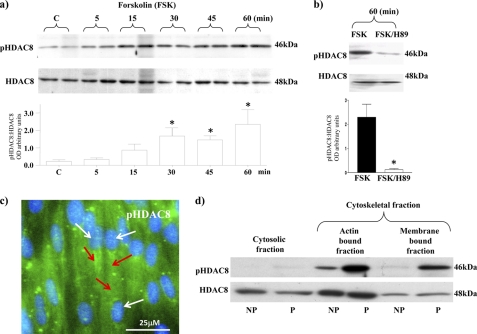
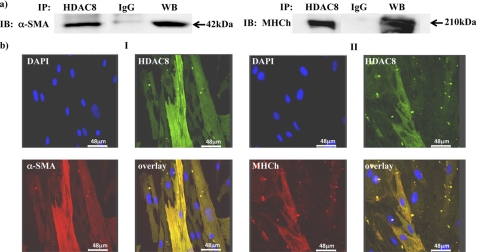
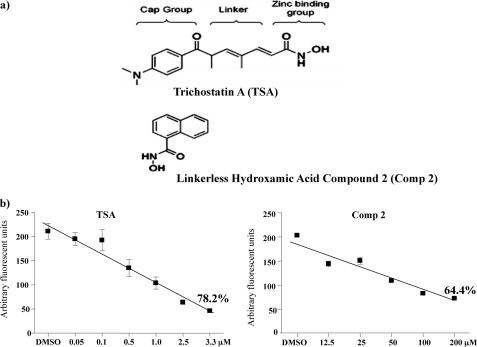
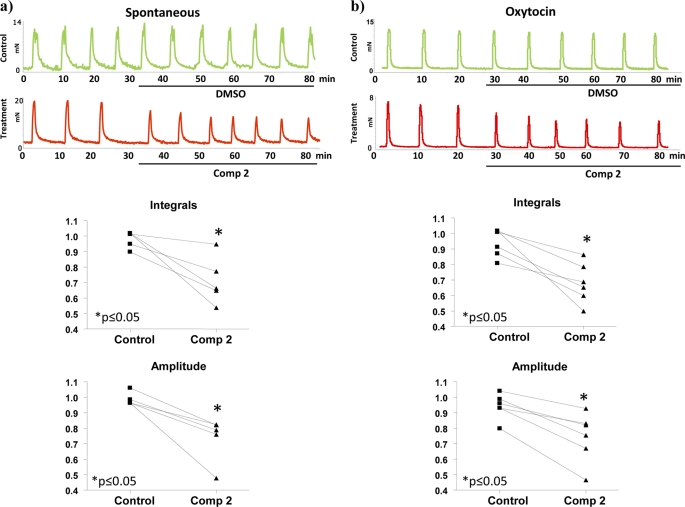
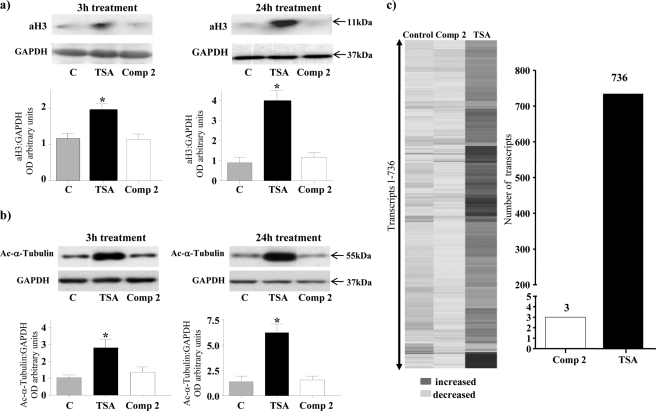
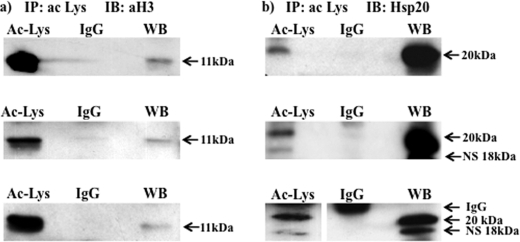
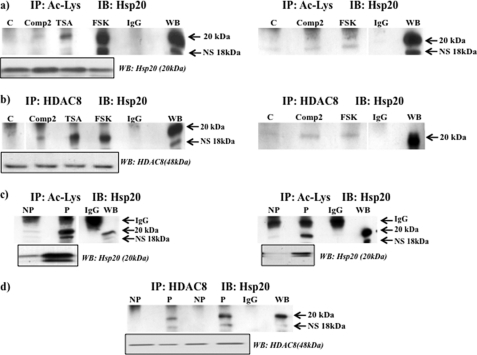
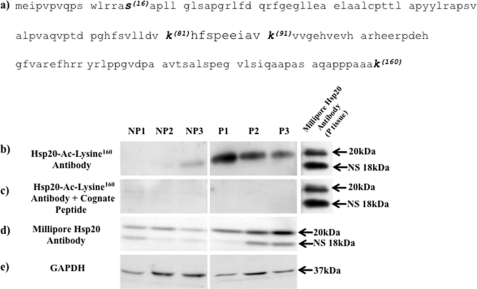
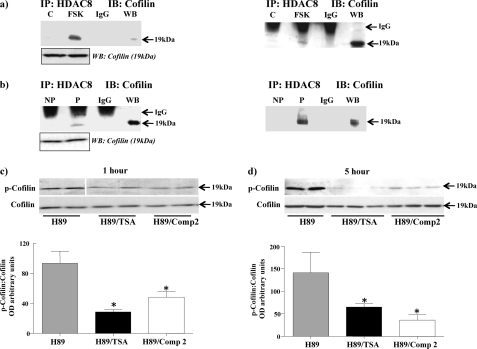
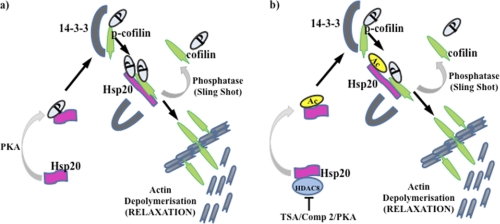
Phosphorylation of heat shock protein 20 (Hsp20) by protein kinase A (PKA) is now recognized as an important regulatory mechanism modulating contractile activity in the human myometrium. Thus agonists that stimulate cyclic AMP production may cause relaxation with resultant beneficial effects on pathologies that affect this tissue such as the onset of premature contractions prior to term. Here we describe for the first time that acetylation of Hsp20 is also a potent post-translational modification that can affect human myometrial activity. We show that histone deacetylase 8 (HDAC8) is a non-nuclear lysine deacetylase (KDAC) that can interact with Hsp20 to affect its acetylation. Importantly, use of a selective linkerless hydroxamic acid HDAC8 inhibitor increases Hsp20 acetylation with no elevation of nuclear-resident histone acetylation nor marked global gene expression changes. These effects are associated with significant inhibition of spontaneous and oxytocin-augmented contractions of ex vivo human myometrial tissue strips. A potential molecular mechanism by which Hsp20 acetylation can affect myometrial activity by liberating cofilin is described and further high-lights the use of specific effectors of KDACs as therapeutic agents in regulating contractility in this smooth muscle.
Figures
References
-
- Moynihan A. T., Hehir M. P., Sharkey A. M., Robson S. C., Europe-Finner G. N., Morrison J. J. (2008) Am. J. Obstet Gynecol 199, 167.e1–7 - PubMed
-
- Kim S. C., Sprung R., Chen Y., Xu Y., Ball H., Pei J., Cheng T., Kho Y., Xiao H., Xiao L., Grishin N. V., White M., Yang X. J., Zhao Y. (2006) Mol. Cell 23, 607–618 - PubMed
-
- Choudhary C., Kumar C., Gnad F., Nielsen M. L., Rehman M., Walther T. C., Olsen J. V., Mann M. (2009) Science 325, 834–840 - PubMed
-
- MacIntyre D. A., Tyson E. K., Read M., Smith R., Yeo G., Kwek K., Chan E. C. (2008) Endocrinology 149, 245–252 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
