

The pentose moiety of adenosine and inosine is an important energy source for the fermented-meat starter culture Lactobacillus sakei CTC 494
- PMID: 21803903
- PMCID: PMC3187176
- DOI: 10.1128/AEM.00498-11
The pentose moiety of adenosine and inosine is an important energy source for the fermented-meat starter culture Lactobacillus sakei CTC 494
Abstract
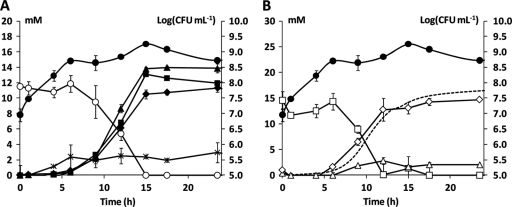
The genome sequence of Lactobacillus sakei 23K has revealed that the species L. sakei harbors several genes involved in the catabolism of energy sources other than glucose in meat, such as glycerol, arginine, and nucleosides. In this study, a screening of 15 L. sakei strains revealed that arginine, inosine, and adenosine could be used as energy sources by all strains. However, no glycerol catabolism occurred in any of the L. sakei strains tested. A detailed kinetic analysis of inosine and adenosine catabolism in the presence of arginine by L. sakei CTC 494, a fermented-meat starter culture, was performed. It showed that nucleoside catabolism occurred as a mixed-acid fermentation in a pH range (pH 5.0 to 6.5) relevant for sausage fermentation. This resulted in the production of a mixture of acetic acid, formic acid, and ethanol from ribose, while the nucleobase (hypoxanthine and adenine in the case of fermentations with inosine and adenosine, respectively) was excreted into the medium stoichiometrically. This indicates that adenosine deaminase activity did not take place. The ratios of the different fermentation end products did not vary with environmental pH, except for the fermentation with inosine at pH 5.0, where lactic acid was produced too. In all cases, no other carbon-containing metabolites were found; carbon dioxide was derived only from arginine catabolism. Arginine was cometabolized in all cases and resulted in the production of both citrulline and ornithine. Based on these results, a pathway for inosine and adenosine catabolism in L. sakei CTC 494 was presented, whereby both nucleosides are directly converted into their nucleobase and ribose, the latter entering the heterolactate pathway. The present study revealed that the pentose moiety (ribose) of the nucleosides inosine and adenosine is an effective fermentable substrate for L. sakei. Thus, the ability to use these energy sources offers a competitive advantage for this species in a meat environment.
Figures






Similar articles
-
The kinetics of the arginine deiminase pathway in the meat starter culture Lactobacillus sakei CTC 494 are pH-dependent.Food Microbiol. 2011 May;28(3):597-604. doi: 10.1016/j.fm.2010.11.016. Epub 2010 Dec 2. Food Microbiol. 2011. PMID: 21356470
-
Comparative genomics of Lactobacillus sakei with emphasis on strains from meat.Mol Genet Genomics. 2011 Apr;285(4):297-311. doi: 10.1007/s00438-011-0608-1. Epub 2011 Mar 3. Mol Genet Genomics. 2011. PMID: 21369871
-
Catabolism of N-acetylneuraminic acid, a fitness function of the food-borne lactic acid bacterium Lactobacillus sakei, involves two newly characterized proteins.Appl Environ Microbiol. 2013 Mar;79(6):2012-8. doi: 10.1128/AEM.03301-12. Epub 2013 Jan 18. Appl Environ Microbiol. 2013. PMID: 23335758 Free PMC article.
-
Lactobacillus sakei: A Starter for Sausage Fermentation, a Protective Culture for Meat Products.Microorganisms. 2017 Sep 6;5(3):56. doi: 10.3390/microorganisms5030056. Microorganisms. 2017. PMID: 28878171 Free PMC article. Review.
-
Latilactobacillus sakei: a candidate probiotic with a key role in food fermentations and health promotion.Crit Rev Food Sci Nutr. 2024;64(4):978-995. doi: 10.1080/10408398.2022.2111402. Epub 2022 Aug 23. Crit Rev Food Sci Nutr. 2024. PMID: 35997270 Review.
Cited by
-
Coagulase-negative Staphylococci favor conversion of arginine into ornithine despite a widespread genetic potential for nitric oxide synthase activity.Appl Environ Microbiol. 2014 Dec;80(24):7741-51. doi: 10.1128/AEM.02298-14. Epub 2014 Oct 3. Appl Environ Microbiol. 2014. PMID: 25281381 Free PMC article.
-
Expression of the arginine deiminase pathway genes in Lactobacillus sakei is strain dependent and is affected by the environmental pH.Appl Environ Microbiol. 2012 Jul;78(14):4874-83. doi: 10.1128/AEM.07724-11. Epub 2012 Apr 27. Appl Environ Microbiol. 2012. PMID: 22544250 Free PMC article.
-
Competition between Starter Cultures and Wild Microbial Population in Sausage Fermentation: A Case Study Regarding a Typical Italian Salami (Ventricina).Foods. 2021 Sep 10;10(9):2138. doi: 10.3390/foods10092138. Foods. 2021. PMID: 34574248 Free PMC article.
-
Contribution of omics to biopreservation: Toward food microbiome engineering.Front Microbiol. 2022 Aug 2;13:951182. doi: 10.3389/fmicb.2022.951182. eCollection 2022. Front Microbiol. 2022. PMID: 35983334 Free PMC article. Review.
-
Shotgun Metagenomics and Volatilome Profile of the Microbiota of Fermented Sausages.Appl Environ Microbiol. 2018 Jan 17;84(3):e02120-17. doi: 10.1128/AEM.02120-17. Print 2018 Feb 1. Appl Environ Microbiol. 2018. PMID: 29196291 Free PMC article.
References
-
- Belitz H. D., Grosch W., Schieberle P. (ed.). 2004. Meat, p. 566–618. In Food Chemistry. Springer, New York, NY
-
- Chaillou S., et al. 2005. The complete genome sequence of the meat-borne lactic acid bacterium Lactobacillus sakei 23K. Nat. Biotechnol. 23: 1527–1533 - PubMed
-
- Champomier-Vergès M. C., Chaillou S., Cornet M., Zagorec M. 2001. Lactobacillus sakei: recent developments and future prospects. Res. Microbiol. 152: 839–848 - PubMed
-
- Champomier Vergès M. C., et al. 1999. Relationships between arginine degradation, pH and survival in Lactobacillus sakei. FEMS Microbiol. Lett. 180: 297–304 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
