

Functional characterization of the sciarid BhC4-1 core promoter in transgenic Drosophila
- PMID: 21806810
- PMCID: PMC3160885
- DOI: 10.1186/1471-2199-12-32
Functional characterization of the sciarid BhC4-1 core promoter in transgenic Drosophila
Abstract
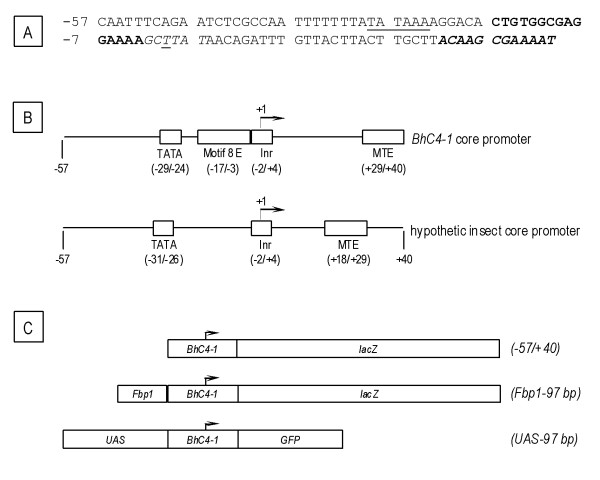
Background: Core promoters are cis-regulatory modules to which bind the basal transcriptional machinery and which participate in the regulation of transcription initiation. Although core promoters have not been extensively investigated through functional assays in a chromosomal context, the available data suggested that the response of a given core promoter might vary depending on the promoter context. Previous studies suggest that a (-57/+40) fragment constitutes the core promoter of the BhC4-1 gene which is located in DNA puff C4 of the sciarid fly Bradysia hygida. Here we tested this (-57/+40) fragment in distinct regulatory contexts in order to verify if promoter context affects its core promoter activity.
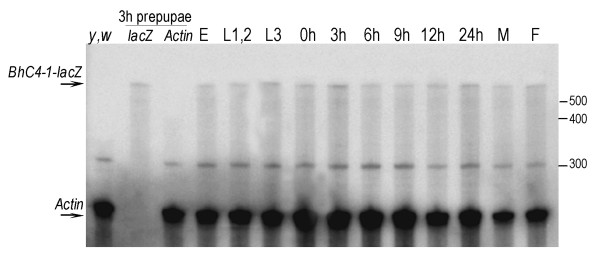
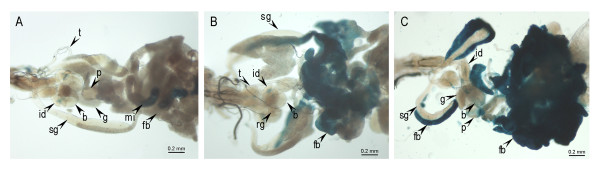
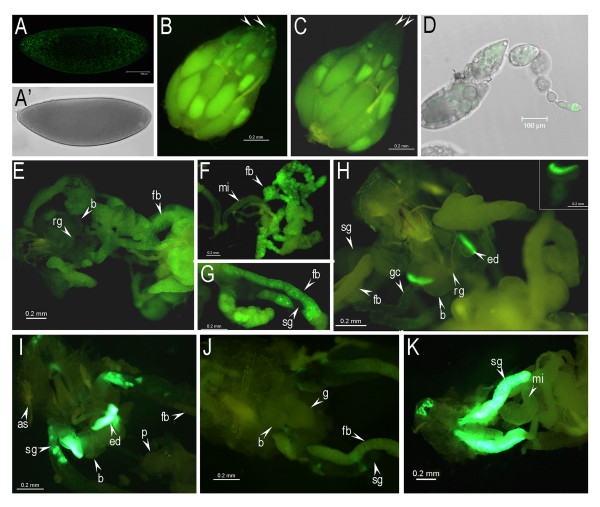
Results: Consistent with the activity of a core promoter, we showed that in the absence of upstream regulatory sequences the (-57/+40) fragment drives low levels of reporter gene mRNA expression throughout development in transgenic Drosophila. By assaying the (-57/+40) fragment in two distinct regulatory contexts, either downstream of the previously characterized Fbp1 enhancer or downstream of the UAS element, we showed that the BhC4-1 core promoter drives regulated transcription in both the germline and in various tissues throughout development. Furthermore, the use of the BhC4-1 core promoter in a UAS construct significantly reduced salivary gland ectopic expression in third instar larvae, which was previously described to occur in the context of the GAL4/UAS system.
Conclusions: Our results from functional analysis in transgenic Drosophila show that the BhC4-1 core promoter drives gene expression regardless of the promoter context that was assayed. New insights into the functioning of the GAL4/UAS system in Drosophila were obtained, indicating that the presence of the SV40 sequence in the 3' UTR of a UAS construct does not preclude expression in the germline. Furthermore, our analysis indicated that ectopic salivary gland expression in the GAL4/UAS system does not depend only on sequences present in the GAL4 construct, but can also be affected by the core promoter sequences in the UAS construct. In this context, we propose that the sciarid BhC4-1 core promoter constitutes a valuable core promoter which can be employed in functional assays in insects.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
