

The historical biogeography of Mammalia
- PMID: 21807730
- PMCID: PMC3138613
- DOI: 10.1098/rstb.2011.0023
The historical biogeography of Mammalia
Abstract
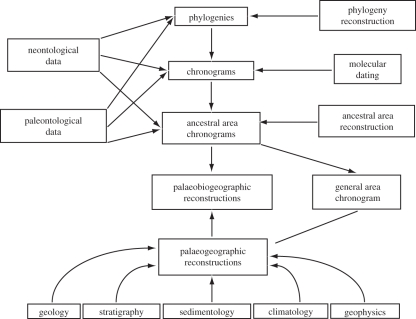
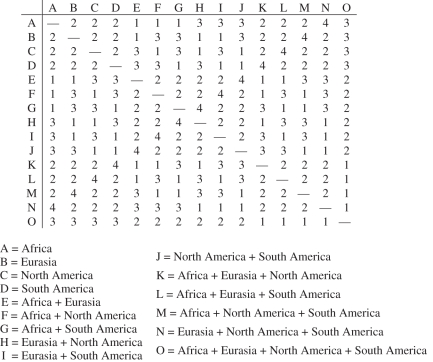
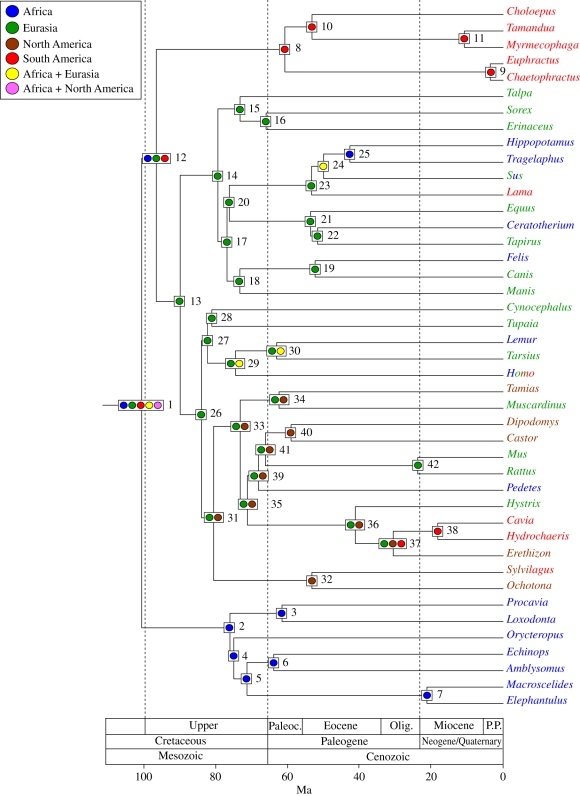
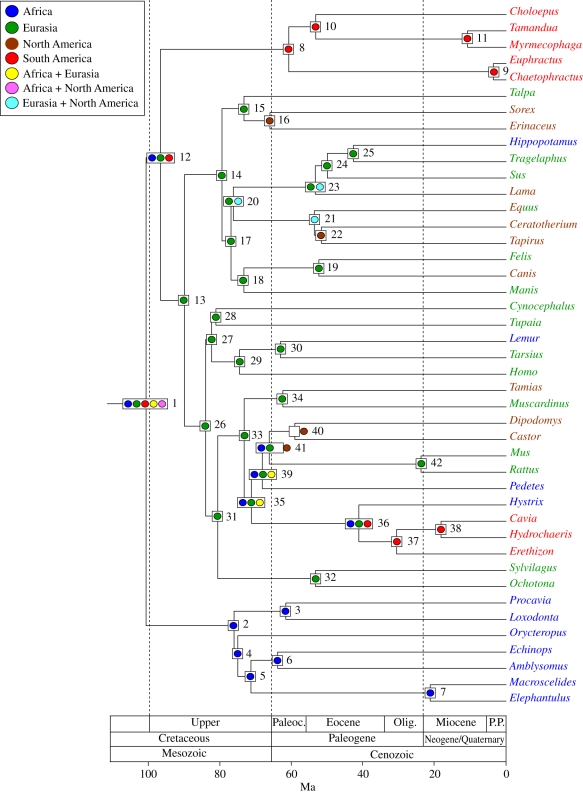
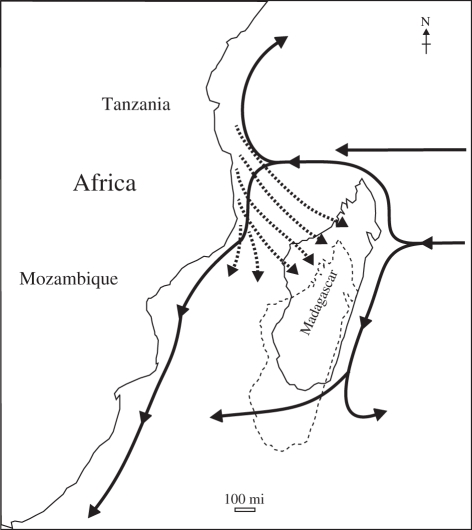
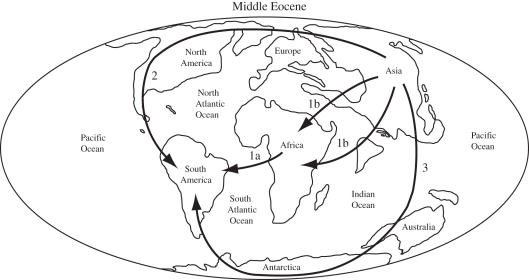
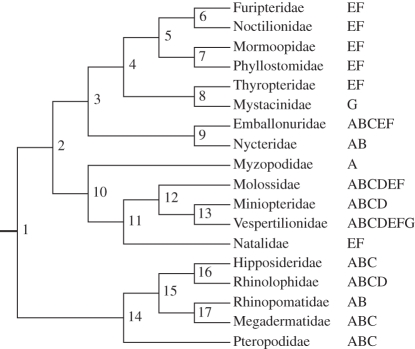
Palaeobiogeographic reconstructions are underpinned by phylogenies, divergence times and ancestral area reconstructions, which together yield ancestral area chronograms that provide a basis for proposing and testing hypotheses of dispersal and vicariance. Methods for area coding include multi-state coding with a single character, binary coding with multiple characters and string coding. Ancestral reconstruction methods are divided into parsimony versus Bayesian/likelihood approaches. We compared nine methods for reconstructing ancestral areas for placental mammals. Ambiguous reconstructions were a problem for all methods. Important differences resulted from coding areas based on the geographical ranges of extant species versus the geographical provenance of the oldest fossil for each lineage. Africa and South America were reconstructed as the ancestral areas for Afrotheria and Xenarthra, respectively. Most methods reconstructed Eurasia as the ancestral area for Boreoeutheria, Euarchontoglires and Laurasiatheria. The coincidence of molecular dates for the separation of Afrotheria and Xenarthra at approximately 100 Ma with the plate tectonic sundering of Africa and South America hints at the importance of vicariance in the early history of Placentalia. Dispersal has also been important including the origins of Madagascar's endemic mammal fauna. Further studies will benefit from increased taxon sampling and the application of new ancestral area reconstruction methods.
Figures
References
-
- Jones K. E., Safi K. 2011. Ecology and evolution of mammalian biodiversity. Phil. Trans. R. Soc. B 366, 2451–2461 10.1098/rstb.2011.0090 (doi:10.1098/rstb.2011.0090) - DOI - PMC - PubMed
-
- Simpson G. G. 1940. Mammals and land bridges. J. Washington D.C. Acad. Sci. 30, 137–163
-
- Springer M. S., Burk-Herrick A., Meredith R., Eizirik E., Teeling E., O'Brien S. J., Murphy W. J. 2007. The adequacy of morphology for reconstructing the early history of placental mammals. Syst. Biol. 56, 673–684 10.1080/10635150701491149 (doi:10.1080/10635150701491149) - DOI - PubMed
-
- Springer M. S., Meredith R. W., Eizirik E., Teeling E., Murphy W. J. 2008. Morphology and placental mammal phylogeny. Syst. Biol. 57, 499–503 10.1080/10635150802164504 (doi:10.1080/10635150802164504) - DOI - PubMed
-
- Bollback J. P. 2006. SIMMAP: stochastic character mapping of discrete traits on phylogenies. BMC Bioinform. 7, 88. 10.1186/1471-2105-7-88 (doi:10.1186/1471-2105-7-88) - DOI - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources