

Angiogenesis
- PMID: 21807843
- PMCID: PMC3140681
- DOI: 10.1101/cshperspect.a005090
Angiogenesis
Abstract
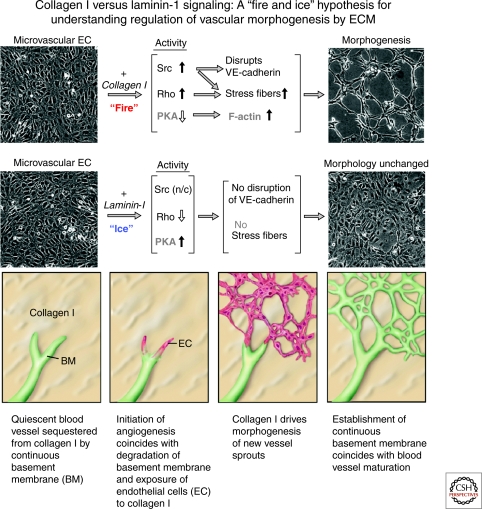
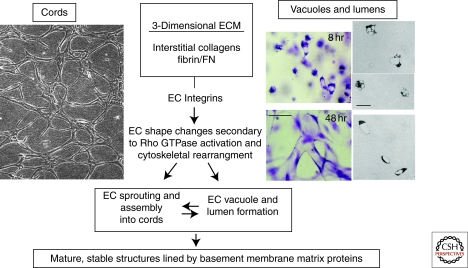
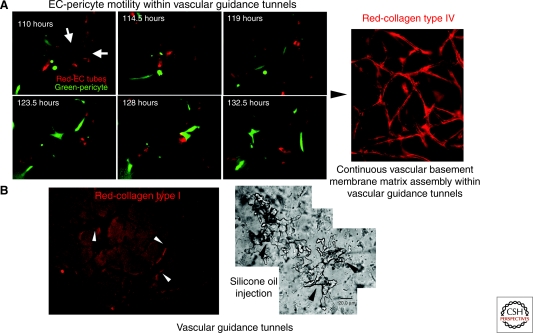
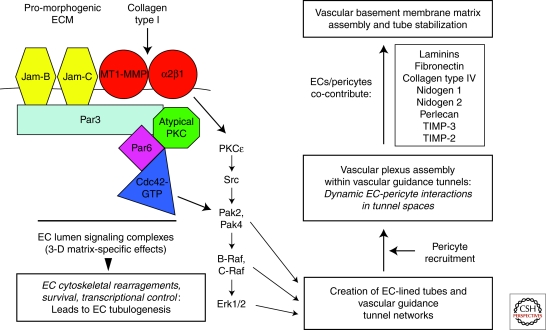
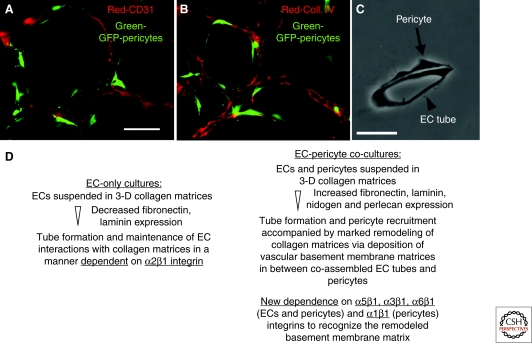
Extracellular matrix (ECM) is essential for all stages of angiogenesis. In the adult, angiogenesis begins with endothelial cell (EC) activation, degradation of vascular basement membrane, and vascular sprouting within interstitial matrix. During this sprouting phase, ECM binding to integrins provides critical signaling support for EC proliferation, survival, and migration. ECM also signals the EC cytoskeleton to initiate blood vessel morphogenesis. Dynamic remodeling of ECM, particularly by membrane-type matrix metalloproteases (MT-MMPs), coordinates formation of vascular tubes with lumens and provides guidance tunnels for pericytes that assist ECs in the assembly of vascular basement membrane. ECM also provides a binding scaffold for a variety of cytokines that exert essential signaling functions during angiogenesis. In the embryo, ECM is equally critical for angiogenesis and vessel stabilization, although there are likely important distinctions from the adult because of differences in composition and abundance of specific ECM components.
Figures
References
-
- Aloisi M, Schiaffino S 1971. Growth of elementary blood vessels in diffusion chambers: Electron microscopy of capillary morphogenesis. Virchows Arch Abt B Zellpath 8: 328–341 - PubMed
-
- Aplin AE, Short SM, Juliano RL 1999. Anchorage-dependent regulation of the mitogen-activated protein kinase cascade by growth factors is supported by a variety of integrin α chains. J Biol Chem 274: 31223–31228 - PubMed
-
- Aplin AC, Fogel E, Zorzi P, Nicosia RF 2008. The aortic ring model of angiogenesis. Meth Enzymol 443: 119–136 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources