

The LYR protein Mzm1 functions in the insertion of the Rieske Fe/S protein in yeast mitochondria
- PMID: 21807901
- PMCID: PMC3187353
- DOI: 10.1128/MCB.05673-11
The LYR protein Mzm1 functions in the insertion of the Rieske Fe/S protein in yeast mitochondria
Abstract
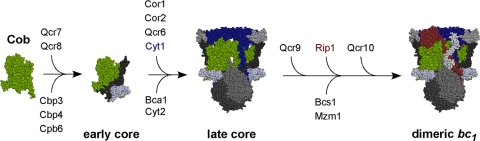
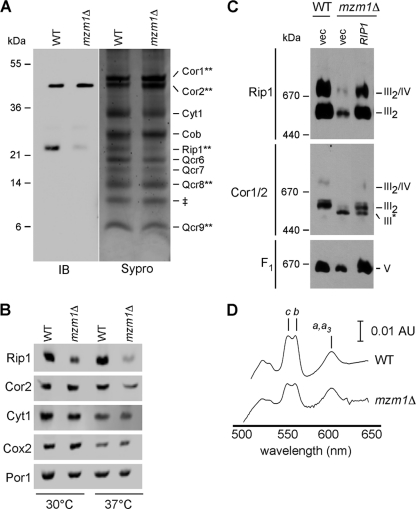
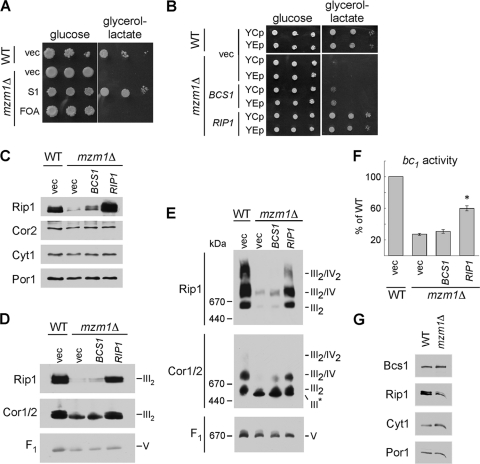
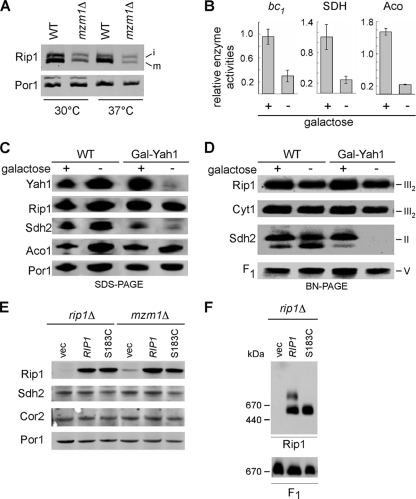
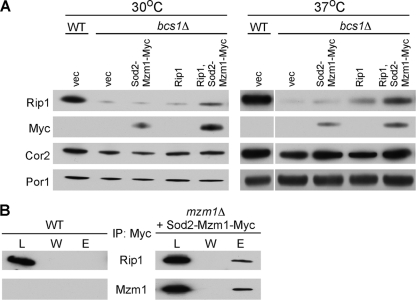
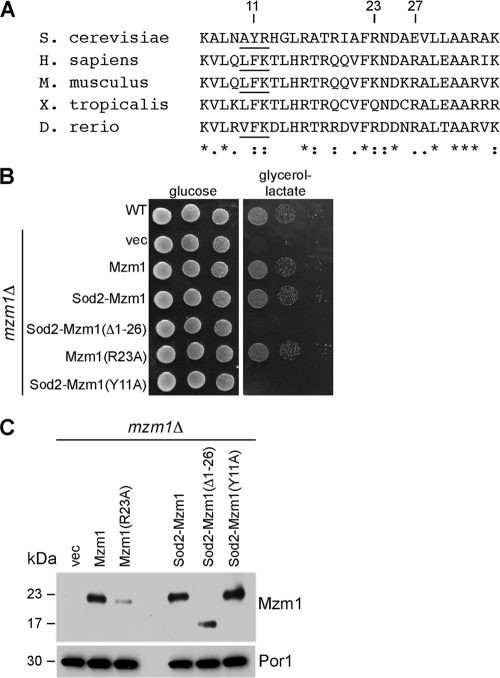
The assembly of the cytochrome bc(1) complex in Saccharomyces cerevisiae is shown to be conditionally dependent on a novel factor, Mzm1. Cells lacking Mzm1 exhibit a modest bc(1) defect at 30°C, but the defect is exacerbated at elevated temperatures. Formation of bc(1) is stalled in mzm1Δ cells at a late assembly intermediate lacking the Rieske iron-sulfur protein Rip1. Rip1 levels are markedly attenuated in mzm1Δ cells at elevated temperatures. Respiratory growth can be restored in the mutant cells by the overexpression of the Rip1 subunit. Elevated levels of Mzm1 enhance the stabilization of Rip1 through physical interaction, suggesting that Mzm1 may be an important Rip1 chaperone especially under heat stress. Mzm1 may function primarily to stabilize Rip1 prior to inner membrane (IM) insertion or alternatively to aid in the presentation of Rip1 to the inner membrane translocation complex for extrusion of the folded domain containing the iron-sulfur center.
Figures
References
-
- Brandt U., Uribe S., Schagger H., Trumpower B. L. 1994. Isolation and characterization of QCR10, the nuclear gene encoding the 8.5-kDa subunit 10 of the Saccharomyces cerevisiae cytochrome bc1 complex. J. Biol. Chem. 269:12947–12953 - PubMed
-
- Crivellone M. D., Wu M. A., Tzagoloff A. 1988. Assembly of the mitochondrial membrane system. Analysis of structural mutants of the yeast coenzyme QH2-cytochrome c reductase complex. J. Biol. Chem. 263:14323–14333 - PubMed
-
- Cruciat C. M., Brunner S., Baumann F., Neupert W., Stuart R. A. 2000. The cytochrome bc1 and cytochrome c oxidase complexes associate to form a single supracomplex in yeast mitochondria. J. Biol. Chem. 275:18093–18098 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous