

The Cdc48 ATPase modulates the interaction between two proteolytic factors Ufd2 and Rad23
- PMID: 21807993
- PMCID: PMC3158229
- DOI: 10.1073/pnas.1104051108
The Cdc48 ATPase modulates the interaction between two proteolytic factors Ufd2 and Rad23
Abstract
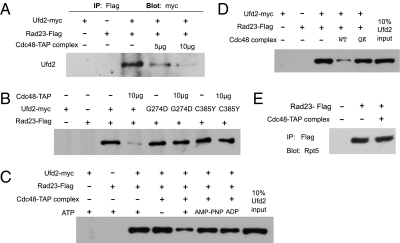
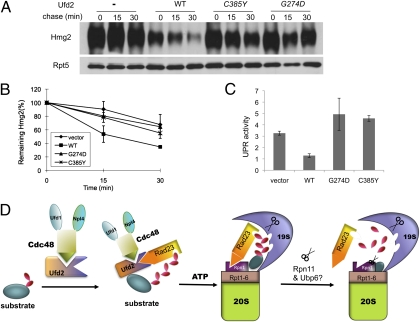
Rad23 and cell division cycle protein 48 (Cdc48), two key regulators of postubiquitylation events, act on distinct and overlapping sets of substrates. The principle underlying their division of labor and cooperation in proteolysis remains elusive. Both Rad23 and Cdc48 bind a ubiquitin protein ligase ubiquitin fusion degradation-2 (Ufd2), and regulate the degradation of Ufd2 substrates. With its ability to bind ubiquitin chains directly and the proteasome via different domains, Rad23 serves as a bridge linking ubiquitylated substrates to the proteasome. The significance and specific role of the Ufd2-Cdc48 interaction are unclear. Here, we demonstrate that mutations in Ufd2 alter its interaction with Cdc48 and impair its function in substrate proteolysis but not in ubiquitylation. Furthermore, Cdc48 promotes the disassembly of the Ufd2-Rad23 complex in an manner that is dependent on ATP and Ufd2 binding, revealing a biochemical role for Cdc48. Rad23 was shown to bind separately to Ufd2 and to the proteasome subunit Rpn1, which define two distinct steps in proteolysis. The action of Cdc48 could free Rad23 from Ufd2 to allow its subsequent association with Rpn1, which in turn may facilitate the orderly transfer of the substrate from the ubiquitylation apparatus to the proteasome.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
