

Social conflict drives the evolutionary divergence of quorum sensing
- PMID: 21807995
- PMCID: PMC3158151
- DOI: 10.1073/pnas.1102923108
Social conflict drives the evolutionary divergence of quorum sensing
Abstract
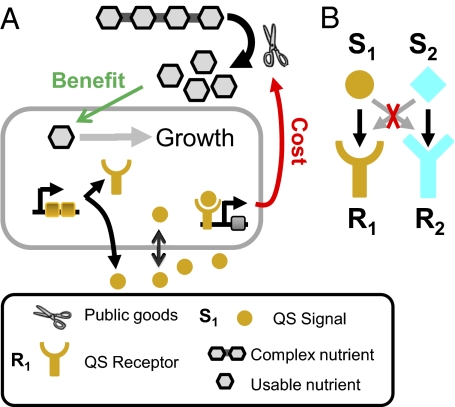
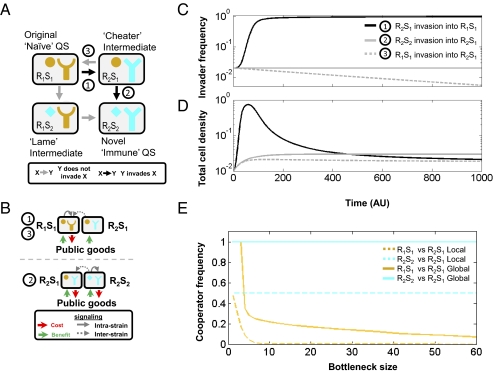
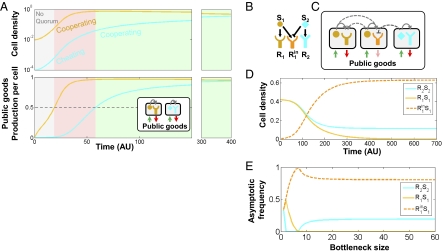
In microbial "quorum sensing" (QS) communication systems, microbes produce and respond to a signaling molecule, enabling a cooperative response at high cell densities. Many species of bacteria show fast, intraspecific, evolutionary divergence of their QS pathway specificity--signaling molecules activate cognate receptors in the same strain but fail to activate, and sometimes inhibit, those of other strains. Despite many molecular studies, it has remained unclear how a signaling molecule and receptor can coevolve, what maintains diversity, and what drives the evolution of cross-inhibition. Here I use mathematical analysis to show that when QS controls the production of extracellular enzymes--"public goods"--diversification can readily evolve. Coevolution is positively selected by cycles of alternating "cheating" receptor mutations and "cheating immunity" signaling mutations. The maintenance of diversity and the evolution of cross-inhibition between strains are facilitated by facultative cheating between the competing strains. My results suggest a role for complex social strategies in the long-term evolution of QS systems. More generally, my model of QS divergence suggests a form of kin recognition where different kin types coexist in unstructured populations.
Conflict of interest statement
The author declares no conflict of interest.
Figures
Similar articles
-
Microbial communication, cooperation and cheating: quorum sensing drives the evolution of cooperation in bacteria.PLoS One. 2009 Aug 17;4(8):e6655. doi: 10.1371/journal.pone.0006655. PLoS One. 2009. PMID: 19684853 Free PMC article.
-
Facultative cheating supports the coexistence of diverse quorum-sensing alleles.Proc Natl Acad Sci U S A. 2016 Feb 23;113(8):2152-7. doi: 10.1073/pnas.1520615113. Epub 2016 Jan 19. Proc Natl Acad Sci U S A. 2016. PMID: 26787913 Free PMC article.
-
Quorum Sensing Protects Pseudomonas aeruginosa against Cheating by Other Species in a Laboratory Coculture Model.J Bacteriol. 2015 Oct;197(19):3154-9. doi: 10.1128/JB.00482-15. Epub 2015 Jul 20. J Bacteriol. 2015. PMID: 26195596 Free PMC article.
-
Evolutionary theory of bacterial quorum sensing: when is a signal not a signal?Philos Trans R Soc Lond B Biol Sci. 2007 Jul 29;362(1483):1241-9. doi: 10.1098/rstb.2007.2049. Philos Trans R Soc Lond B Biol Sci. 2007. PMID: 17360270 Free PMC article. Review.
-
Quorum sensing: A less known mode of communication among fungi.Microbiol Res. 2018 May;210:51-58. doi: 10.1016/j.micres.2018.03.007. Epub 2018 Mar 21. Microbiol Res. 2018. PMID: 29625658 Review.
Cited by
-
Black Queen Hypothesis, partial privatization, and quorum sensing evolution.PLoS One. 2022 Nov 30;17(11):e0278449. doi: 10.1371/journal.pone.0278449. eCollection 2022. PLoS One. 2022. PMID: 36449503 Free PMC article.
-
Detecting the molecular signature of social conflict: theory and a test with bacterial quorum sensing genes.Am Nat. 2012 Apr;179(4):436-50. doi: 10.1086/664609. Epub 2012 Mar 6. Am Nat. 2012. PMID: 22437174 Free PMC article.
-
Targeting virulence: can we make evolution-proof drugs?Nat Rev Microbiol. 2014 Apr;12(4):300-8. doi: 10.1038/nrmicro3232. Nat Rev Microbiol. 2014. PMID: 24625893 Review.
-
Evolution of new regulatory functions on biophysically realistic fitness landscapes.Nat Commun. 2017 Aug 9;8(1):216. doi: 10.1038/s41467-017-00238-8. Nat Commun. 2017. PMID: 28790313 Free PMC article.
-
Kin Recognition in Bacteria.Annu Rev Microbiol. 2016 Sep 8;70:143-60. doi: 10.1146/annurev-micro-102215-095325. Epub 2016 Jun 17. Annu Rev Microbiol. 2016. PMID: 27359217 Free PMC article. Review.
References
-
- Waters CM, Bassler BL. Quorum sensing: Cell-to-cell communication in bacteria. Annu Rev Cell Dev Biol. 2005;21:319–346. - PubMed
-
- Ji G, Beavis R, Novick RP. Bacterial interference caused by autoinducing peptide variants. Science. 1997;276:2027–2030. - PubMed
-
- Tran L-SP, Nagai T, Itoh Y. Divergent structure of the ComQXPA quorum-sensing components: Molecular basis of strain-specific communication mechanism in Bacillus subtilis. Mol Microbiol. 2000;37:1159–1171. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
