

Components of cross-frequency modulation in health and disease
- PMID: 21808609
- PMCID: PMC3139214
- DOI: 10.3389/fnsys.2011.00059
Components of cross-frequency modulation in health and disease
Abstract
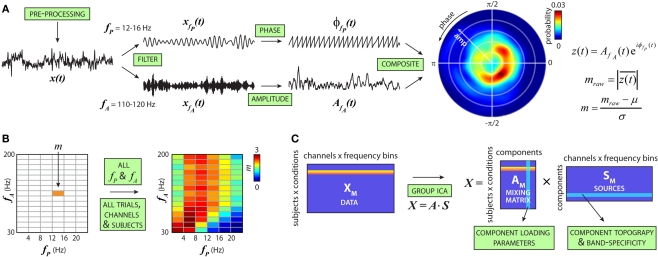
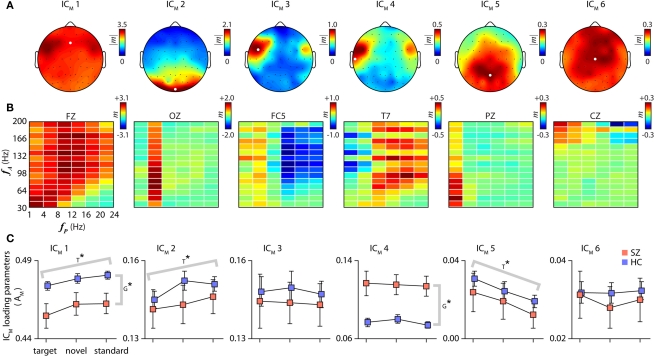
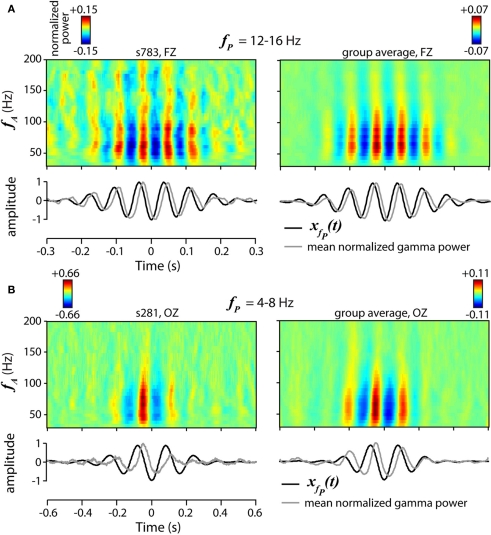
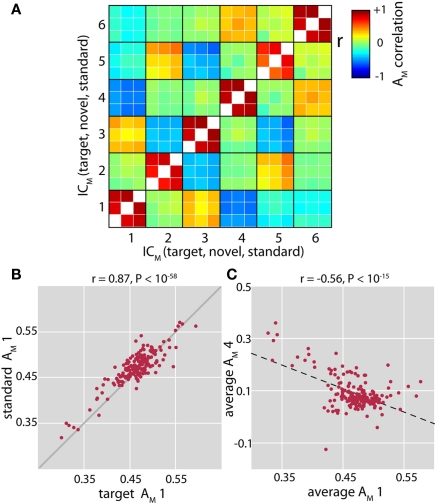
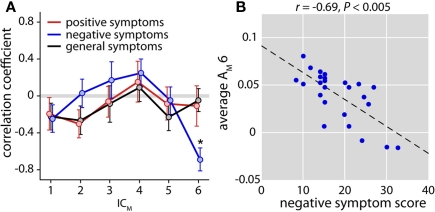
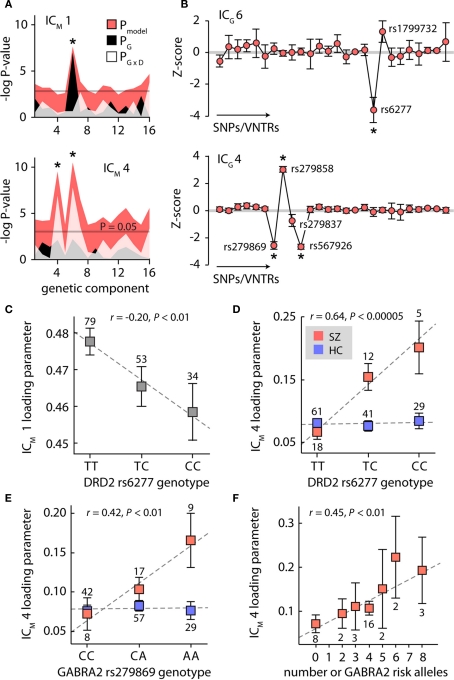
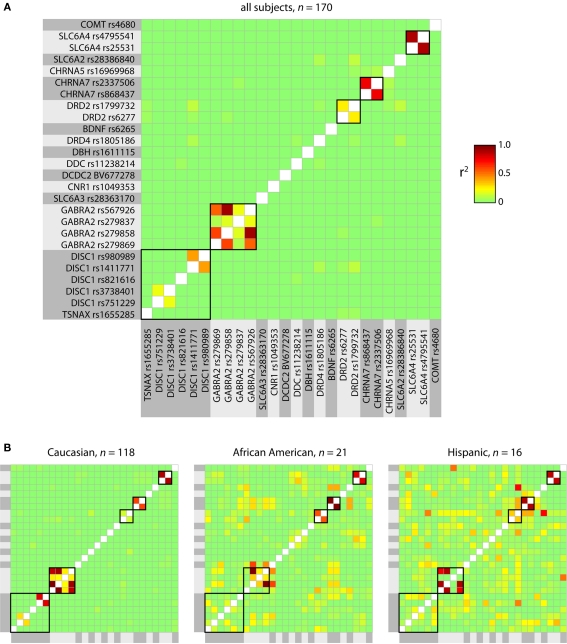
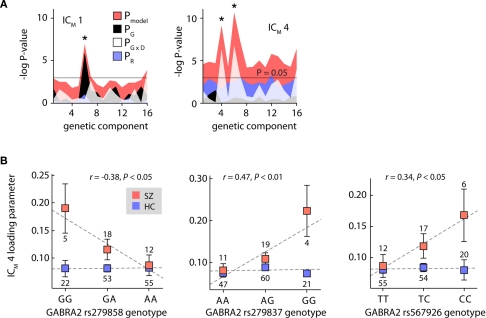
The cognitive deficits associated with schizophrenia are commonly believed to arise from the abnormal temporal integration of information, however a quantitative approach to assess network coordination is lacking. Here, we propose to use cross-frequency modulation (cfM), the dependence of local high-frequency activity on the phase of widespread low-frequency oscillations, as an indicator of network coordination and functional integration. In an exploratory analysis based on pre-existing data, we measured cfM from multi-channel EEG recordings acquired while schizophrenia patients (n = 47) and healthy controls (n = 130) performed an auditory oddball task. Novel application of independent component analysis (ICA) to modulation data delineated components with specific spatial and spectral profiles, the weights of which showed covariation with diagnosis. Global cfM was significantly greater in healthy controls (F(1,175) = 9.25, P < 0.005), while modulation at fronto-temporal electrodes was greater in patients (F(1,175) = 17.5, P < 0.0001). We further found that the weights of schizophrenia-relevant components were associated with genetic polymorphisms at previously identified risk loci. Global cfM decreased with copies of 957C allele in the gene for the dopamine D2 receptor (r = -0.20, P < 0.01) across all subjects. Additionally, greater "aberrant" fronto-temporal modulation in schizophrenia patients was correlated with several polymorphisms in the gene for the α2-subunit of the GABA(A) receptor (GABRA2) as well as the total number of risk alleles in GABRA2 (r = 0.45, P < 0.01). Overall, our results indicate great promise for this approach in establishing patterns of cfM in health and disease and elucidating the roles of oscillatory interactions in functional connectivity.
Keywords: EEG; biomarker; cross-frequency coupling; cross-frequency modulation; independent component analysis; oscillations; schizophrenia.
Figures
References
-
- Andreasen N., Paradiso S., O'Leary D. (1998). “Cognitive dysmetria” as an integrative theory of schizophrenia: a dysfunction in cortical-subcortical-cerebellar circuitry? Schizophr. Bull. 24, 203. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous