

Identification and characterization of DGA2, an acyltransferase of the DGAT1 acyl-CoA:diacylglycerol acyltransferase family in the oleaginous yeast Yarrowia lipolytica. New insights into the storage lipid metabolism of oleaginous yeasts
- PMID: 21808970
- PMCID: PMC3275733
- DOI: 10.1007/s00253-011-3506-x
Identification and characterization of DGA2, an acyltransferase of the DGAT1 acyl-CoA:diacylglycerol acyltransferase family in the oleaginous yeast Yarrowia lipolytica. New insights into the storage lipid metabolism of oleaginous yeasts
Abstract
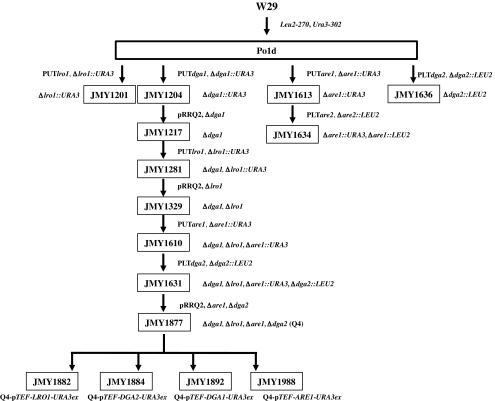
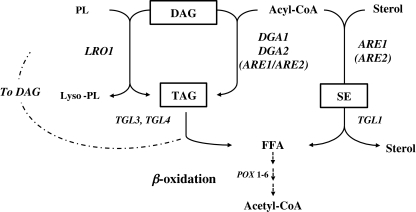
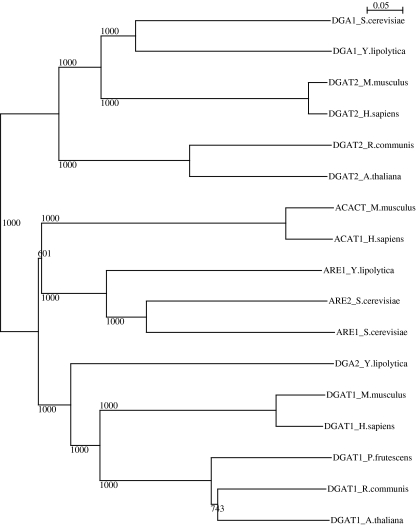
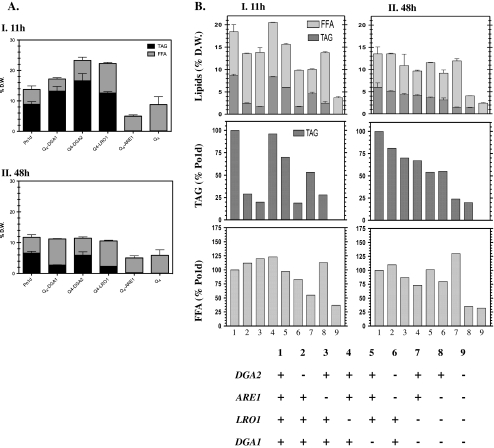
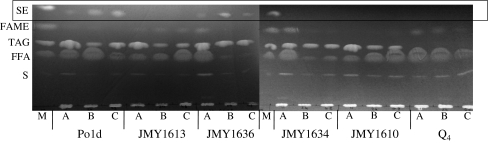
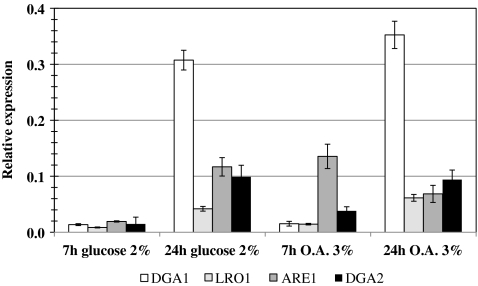
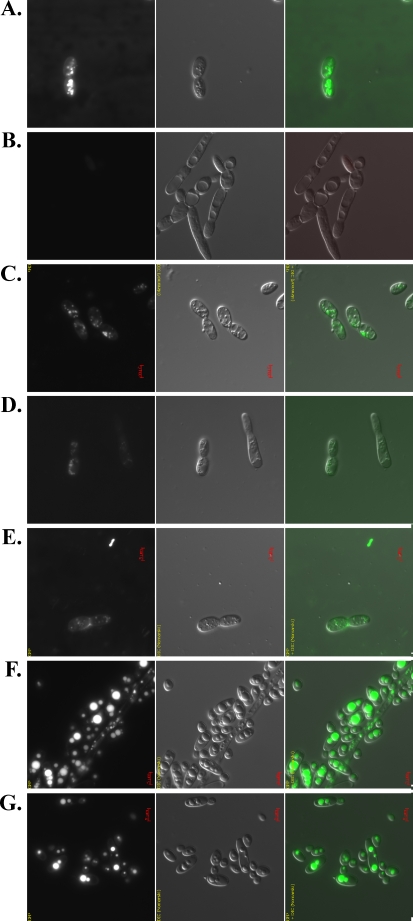
Triacylglycerols (TAG) and steryl esters (SE) are the principal storage lipids in all eukaryotic cells. In yeasts, these storage lipids accumulate within special organelles known as lipid bodies (LB). In the lipid accumulation-oriented metabolism of the oleaginous yeast Yarrowia lipolytica, storage lipids are mostly found in the form of TAG, and only small amounts of SE accumulate. We report here the identification of a new DAG acyltransferase gene, DGA2, homologous to the ARE genes of Saccharomyces cerevisiae. This gene encodes a member of the type 1 acyl-CoA:diacylglycerol acyltransferase family (DGAT1), which has not previously been identified in yeasts, but is commonly found in mammals and plants. Unlike the Are proteins in S. cerevisiae, Dga2p makes a major contribution to TAG synthesis via an acyl-CoA-dependent mechanism and is not involved in SE synthesis. This enzyme appears to affect the size and morphology of LB, suggesting a direct role of storage lipid proteins in LB formation. We report that the Are1p of Y. lipolytica was essential for sterol esterification, as deletion of the encoding gene (ARE1) completely abolished SE synthesis. Unlike its homologs in yeasts, YlARE1 has no DAG acyltransferase activity. We also reconsider the role and function of all four acyltransferase enzymes involved in the final step of neutral lipid synthesis in this oleaginous yeast.
Figures
References
-
- Barth G, Gaillardin C. Yarrowia lipolytica. In: Wolf K, editor. Non conventional yeasts in biotechnology, a handbook. Berlin: Springer; 1996. pp. 313–388.
-
- Barth G, JM B, Dominguez A, Kerscher S, Ogrydziak D, Titorenko V, Gaillardin C. Functional genetics of Yarrowia lipolytica. In: dW JH, editor. Functional genetics of industrial yeasts, vol 1. Topics in current genetics. Berlin: Springer; 2003. pp. 227–271.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
