

A protective antiarrhythmic role of ursodeoxycholic acid in an in vitro rat model of the cholestatic fetal heart
- PMID: 21809354
- PMCID: PMC4900448
- DOI: 10.1002/hep.24492
A protective antiarrhythmic role of ursodeoxycholic acid in an in vitro rat model of the cholestatic fetal heart
Abstract
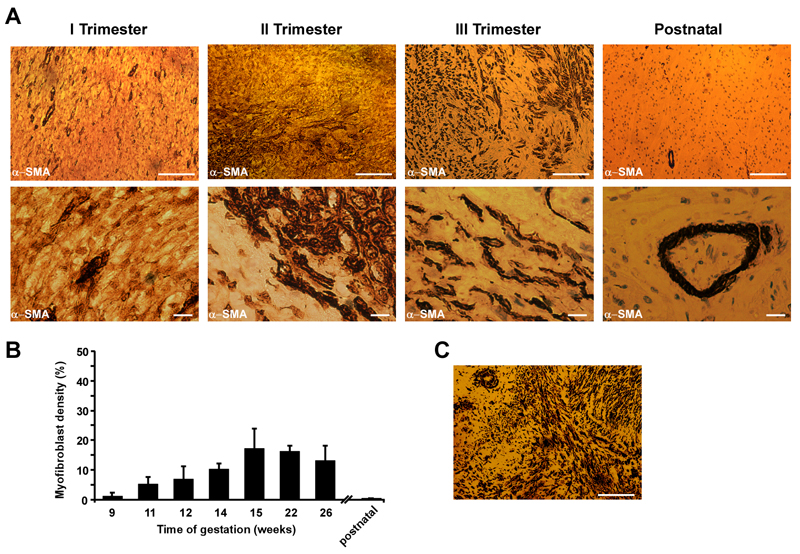
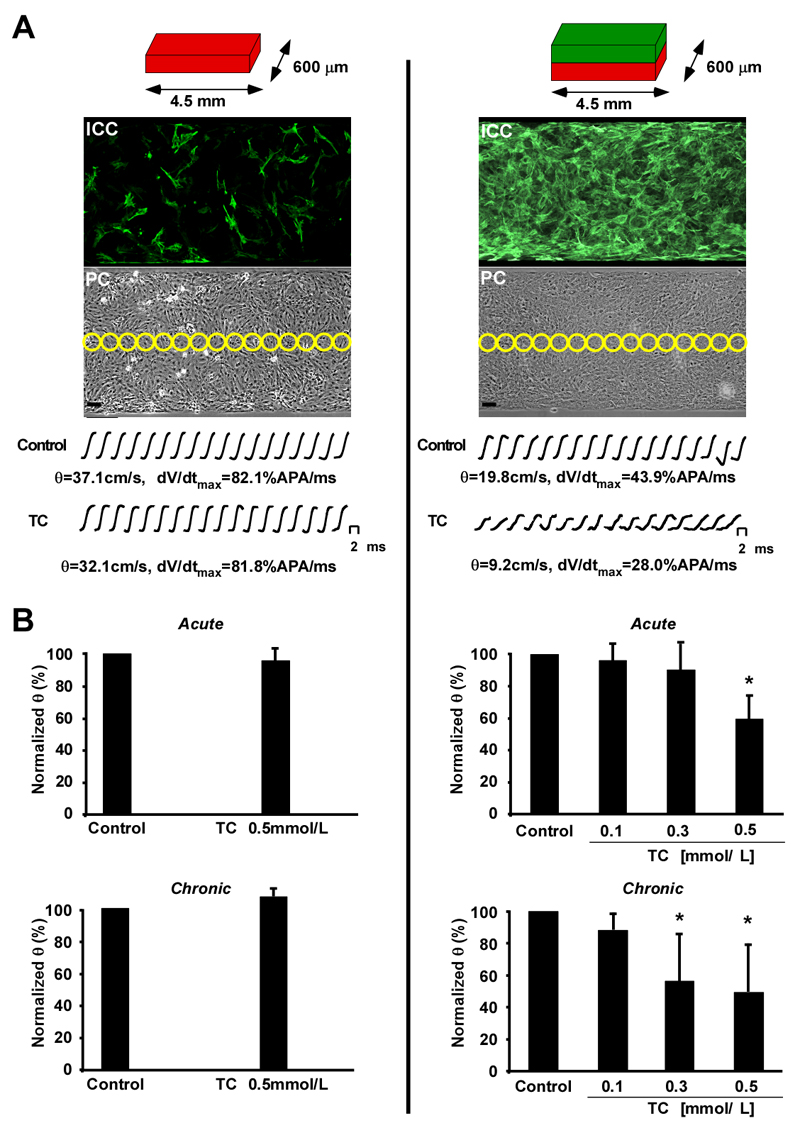
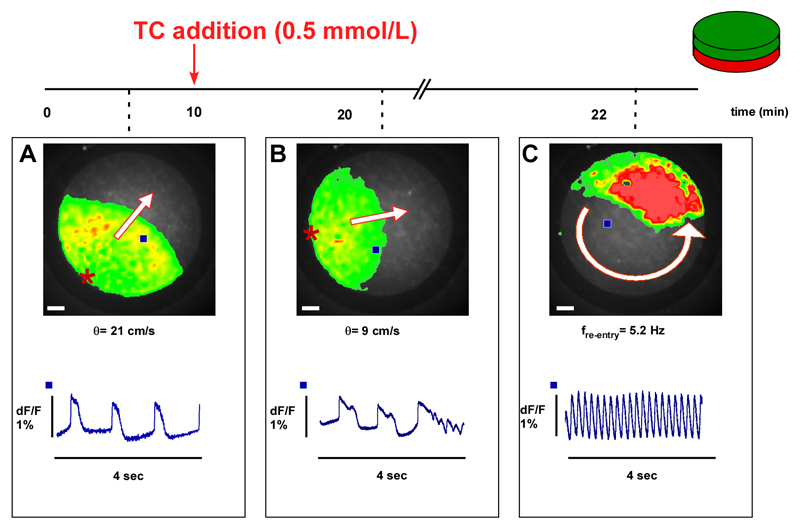
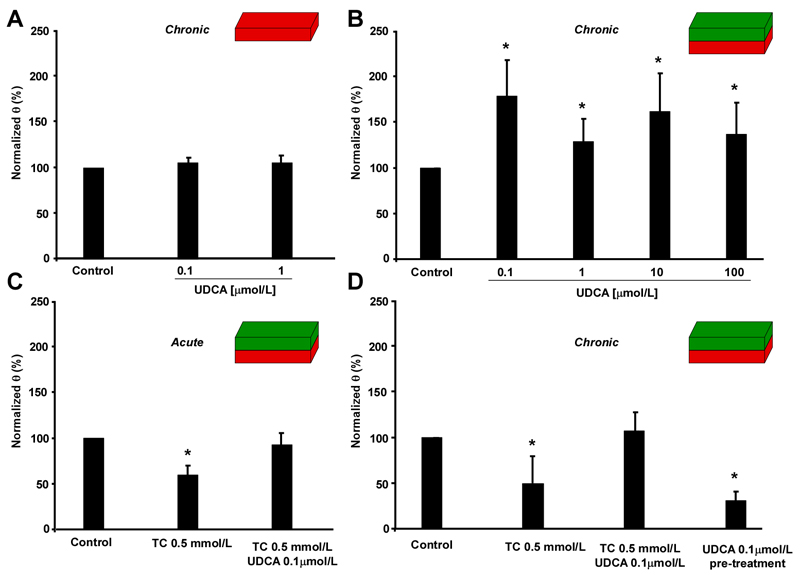
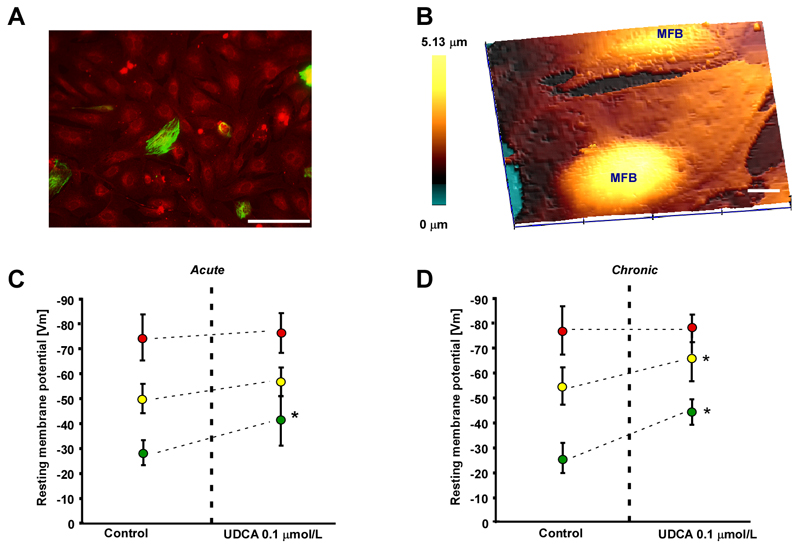
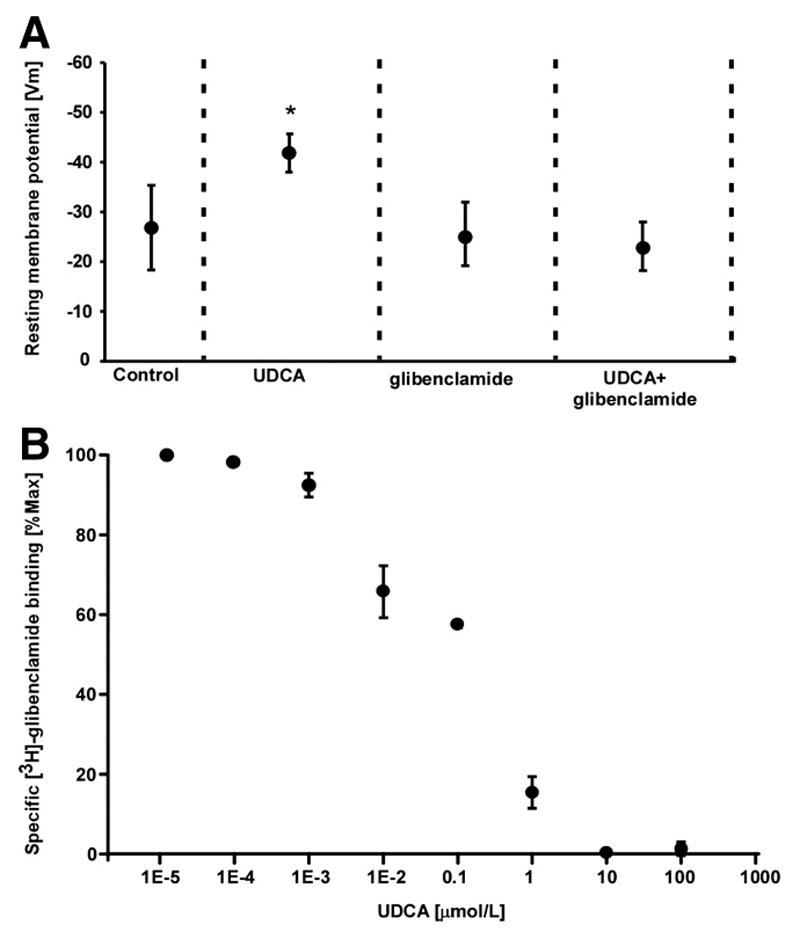
Intrahepatic cholestasis of pregnancy may be complicated by fetal arrhythmia, fetal hypoxia, preterm labor, and, in severe cases, intrauterine death. The precise etiology of fetal death is not known. However, taurocholate has been demonstrated to cause arrhythmia and abnormal calcium dynamics in cardiomyocytes. To identify the underlying reason for increased susceptibility of fetal cardiomyocytes to arrhythmia, we studied myofibroblasts (MFBs), which appear during structural remodeling of the adult diseased heart. In vitro, they depolarize rat cardiomyocytes via heterocellular gap junctional coupling. Recently, it has been hypothesized that ventricular MFBs might appear in the developing human heart, triggered by physiological fetal hypoxia. However, their presence in the fetal heart (FH) and their proarrhythmogenic effects have not been systematically characterized. Immunohistochemistry demonstrated that ventricular MFBs transiently appear in the human FH during gestation. We established two in vitro models of the maternal heart (MH) and FH, both exposed to increasing doses of taurocholate. The MH model consisted of confluent strands of rat cardiomyocytes, whereas for the FH model, we added cardiac MFBs on top of cardiomyocytes. Taurocholate in the FH model, but not in the MH model, slowed conduction velocity from 19 to 9 cm/s, induced early after depolarizations, and resulted in sustained re-entrant arrhythmias. These arrhythmic events were prevented by ursodeoxycholic acid, which hyperpolarized MFB membrane potential by modulating potassium conductance.
Conclusion: These results illustrate that the appearance of MFBs in the FH may contribute to arrhythmias. The above-described mechanism represents a new therapeutic approach for cardiac arrhythmias at the level of MFB.
Copyright © 2011 American Association for the Study of Liver Diseases.
Conflict of interest statement
Figures
References
-
- Glantz A, Marschall HU, Lammert F, Mattsson LA. Intrahepatic cholestasis of pregnancy: a randomized controlled trial comparing dexamethasone and ursodeoxycholic acid. Hepatology. 2005;42:1399–1405. - PubMed
-
- Al Inizi S, Gupta R, Gale A. Fetal tachyarrhythmia with atrial flutter in obstetric cholestasis. Int J Gynaecol Obstet. 2006;93:53–54. - PubMed
-
- Shand AW, Dickinson JE, D'Orsogna L. Refractory fetal supraventricular tachycardia and obstetric cholestasis. Fetal Diagn Ther. 2008;24:277–281. - PubMed
-
- Fisk NM, Storey GN. Fetal outcome in obstetric cholestasis. Br J Obstet Gynaecol. 1988;95:1137–1143. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous