

Functional analysis of all AGAMOUS subfamily members in rice reveals their roles in reproductive organ identity determination and meristem determinacy
- PMID: 21810995
- PMCID: PMC3180796
- DOI: 10.1105/tpc.111.087007
Functional analysis of all AGAMOUS subfamily members in rice reveals their roles in reproductive organ identity determination and meristem determinacy
Abstract
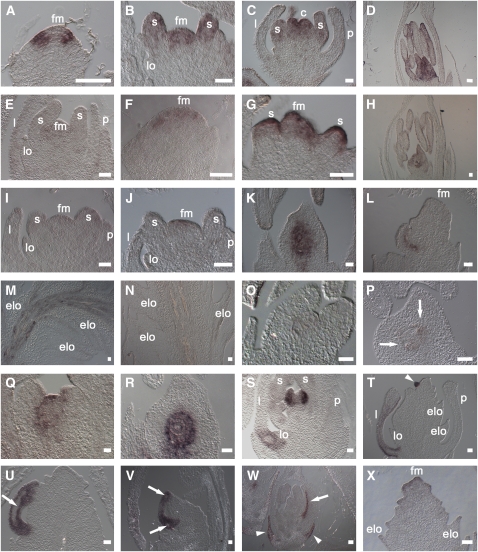
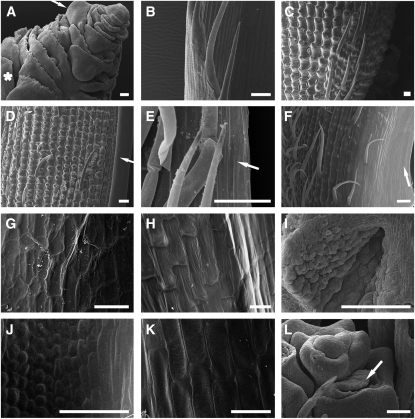
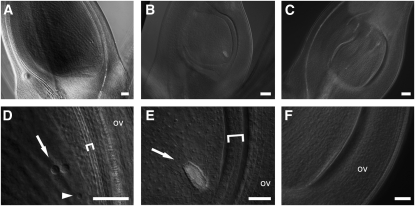
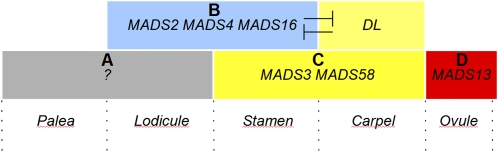
Reproductive organ development is one of the most important steps in the life cycle of plants. Studies using core eudicot species like thale cress (Arabidopsis thaliana) and snapdragon (Antirrhinum majus) have shown that MADS domain transcription factors belonging to the AGAMOUS (AG) subfamily regulate the identity of stamens, carpels, and ovules and that they are important for floral meristem determinacy. Here, we investigate the genetic interactions between the four rice (Oryza sativa) AG subfamily members, MADS3, MADS13, MADS21, and MADS58. Our data show that, in contrast with previous reports, MADS3 and MADS58 determine stamen and carpel identity and, together with MADS13, are important for floral meristem determinacy. In the mads3 mads58 double mutant, we observed a complete loss of reproductive organ identity and massive accumulation of lodicules in the third and fourth floral whorls. MADS21 is an AGL11 lineage gene whose expression is not restricted to ovules. Instead, its expression profile is similar to those of class C genes. However, our genetic analysis shows that MADS21 has no function in stamen, carpel, or ovule identity determination.
Figures

References
-
- Alvarez J., Smyth D.R. (1999). CRABS CLAW and SPATULA, two Arabidopsis genes that control carpel development in parallel with AGAMOUS. Development 126: 2377–2386 - PubMed
-
- Bowman J.L., Smyth D.R. (1999). CRABS CLAW, a gene that regulates carpel and nectary development in Arabidopsis, encodes a novel protein with zinc finger and helix-loop-helix domains. Development 126: 2387–2396 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
