

AlleleSeq: analysis of allele-specific expression and binding in a network framework
- PMID: 21811232
- PMCID: PMC3208341
- DOI: 10.1038/msb.2011.54
AlleleSeq: analysis of allele-specific expression and binding in a network framework
Abstract
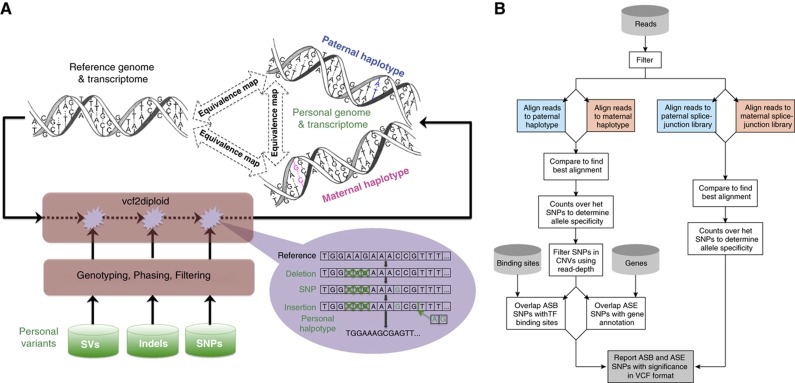
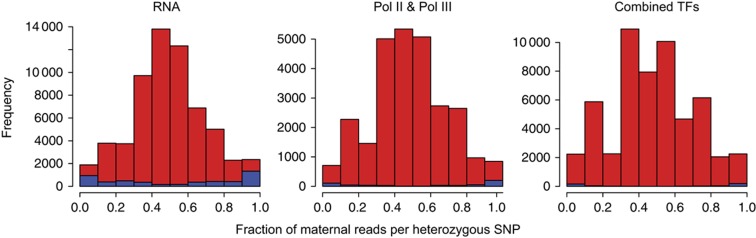
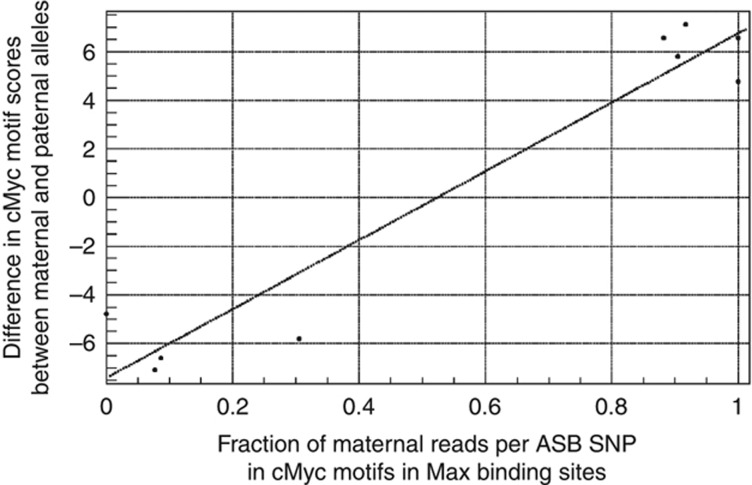
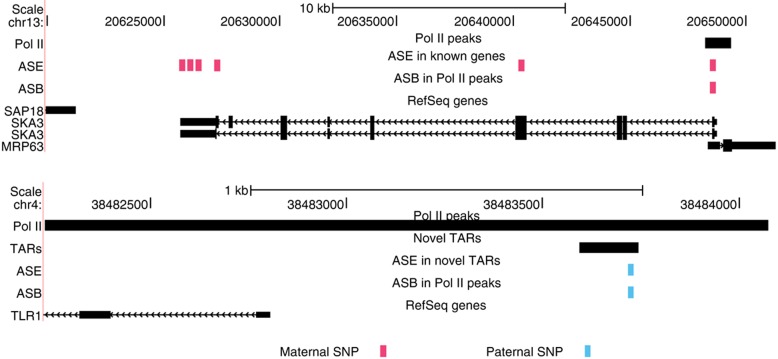
To study allele-specific expression (ASE) and binding (ASB), that is, differences between the maternally and paternally derived alleles, we have developed a computational pipeline (AlleleSeq). Our pipeline initially constructs a diploid personal genome sequence (and corresponding personalized gene annotation) using genomic sequence variants (SNPs, indels, and structural variants), and then identifies allele-specific events with significant differences in the number of mapped reads between maternal and paternal alleles. There are many technical challenges in the construction and alignment of reads to a personal diploid genome sequence that we address, for example, bias of reads mapping to the reference allele. We have applied AlleleSeq to variation data for NA12878 from the 1000 Genomes Project as well as matched, deeply sequenced RNA-Seq and ChIP-Seq data sets generated for this purpose. In addition to observing fairly widespread allele-specific behavior within individual functional genomic data sets (including results consistent with X-chromosome inactivation), we can study the interaction between ASE and ASB. Furthermore, we investigate the coordination between ASE and ASB from multiple transcription factors events using a regulatory network framework. Correlation analyses and network motifs show mostly coordinated ASB and ASE.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
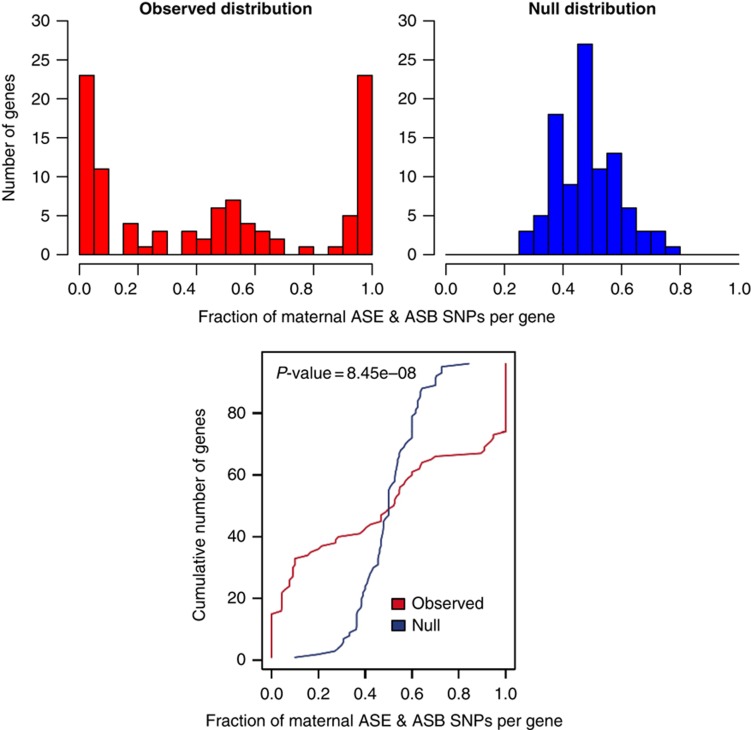
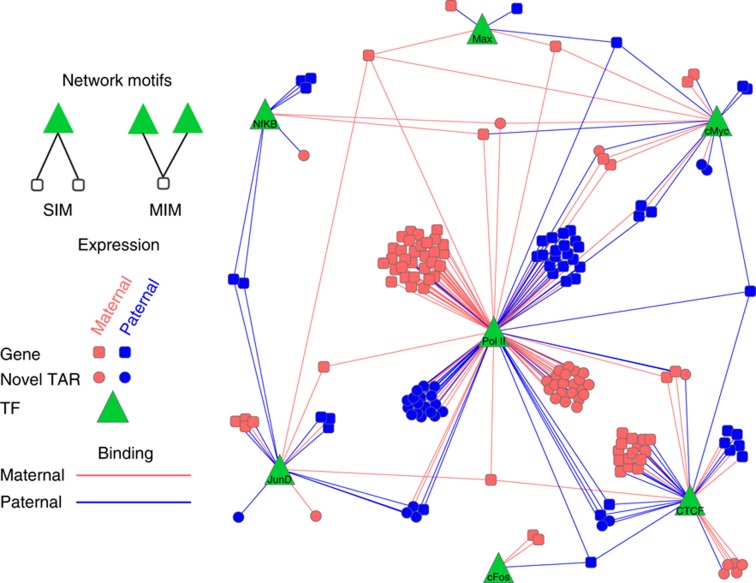
References
-
- Bertone P, Stolc V, Royce TE, Rozowsky JS, Urban AE, Zhu X, Rinn JL, Tongprasit W, Samanta M, Weissman S, Gerstein M, Snyder M (2004) Global identification of human transcribed sequences with genome tiling arrays. Science 306: 2242–2246 - PubMed
-
- Cline MS, Smoot M, Cerami E, Kuchinsky A, Landys N, Workman C, Christmas R, Avila-Campilo I, Creech M, Gross B, Hanspers K, Isserlin R, Kelley R, Killcoyne S, Lotia S, Maere S, Morris J, Ono K, Pavlovic V, Pico AR et al. (2007) Integration of biological networks and gene expression data using Cytoscape. Nat Protoc 2: 2366–2382 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
