

Allele-specific transcriptional elongation regulates monoallelic expression of the IGF2BP1 gene
- PMID: 21812971
- PMCID: PMC3174113
- DOI: 10.1186/1756-8935-4-14
Allele-specific transcriptional elongation regulates monoallelic expression of the IGF2BP1 gene
Abstract
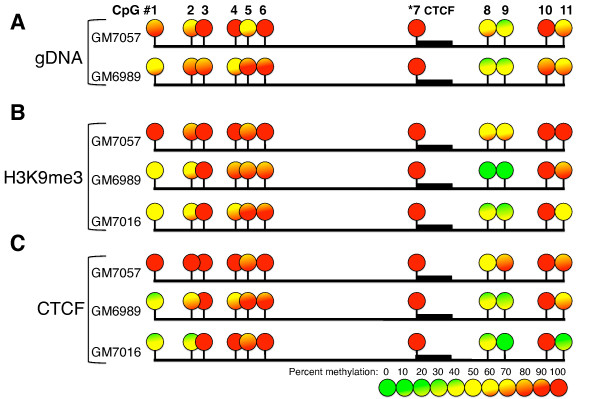
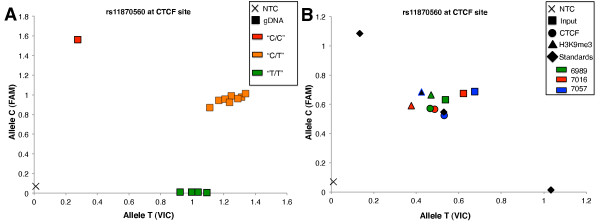
Background: Random monoallelic expression contributes to phenotypic variation of cells and organisms. However, the epigenetic mechanisms by which individual alleles are randomly selected for expression are not known. Taking cues from chromatin signatures at imprinted gene loci such as the insulin-like growth factor 2 gene 2 (IGF2), we evaluated the contribution of CTCF, a zinc finger protein required for parent-of-origin-specific expression of the IGF2 gene, as well as a role for allele-specific association with DNA methylation, histone modification and RNA polymerase II.
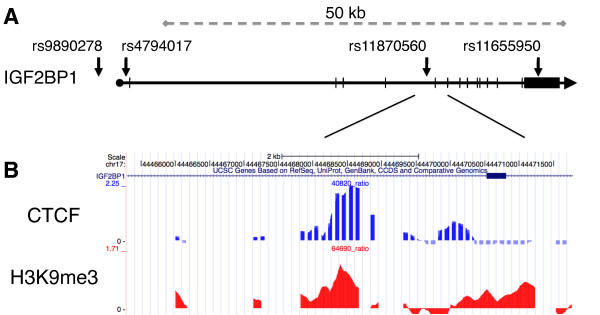
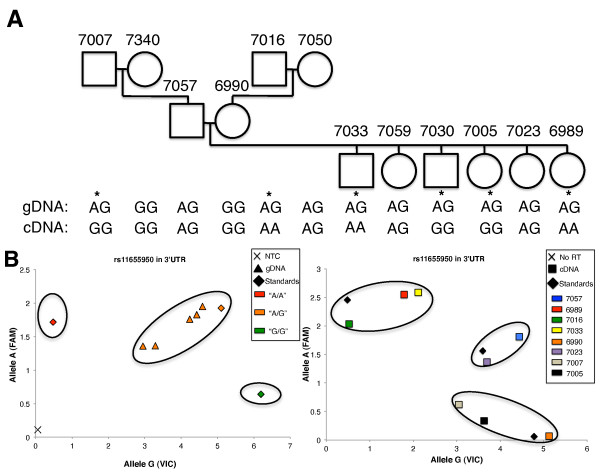
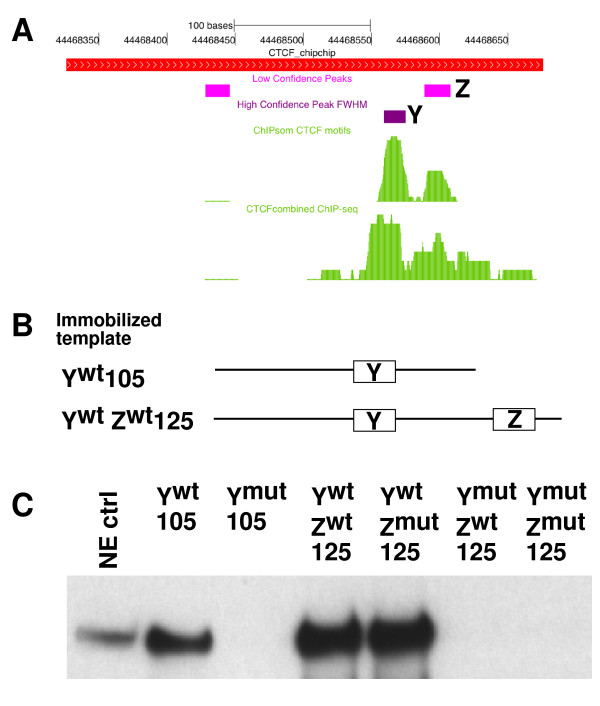
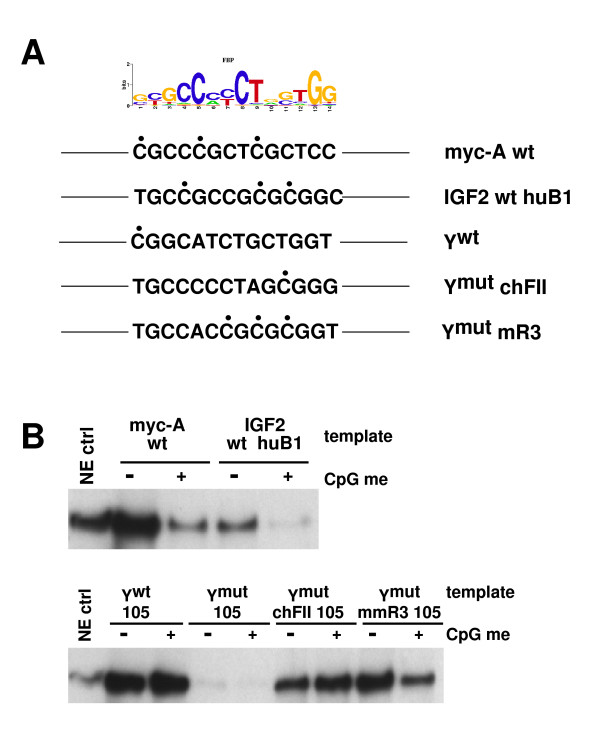
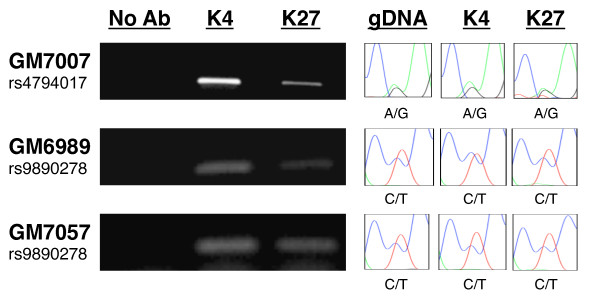
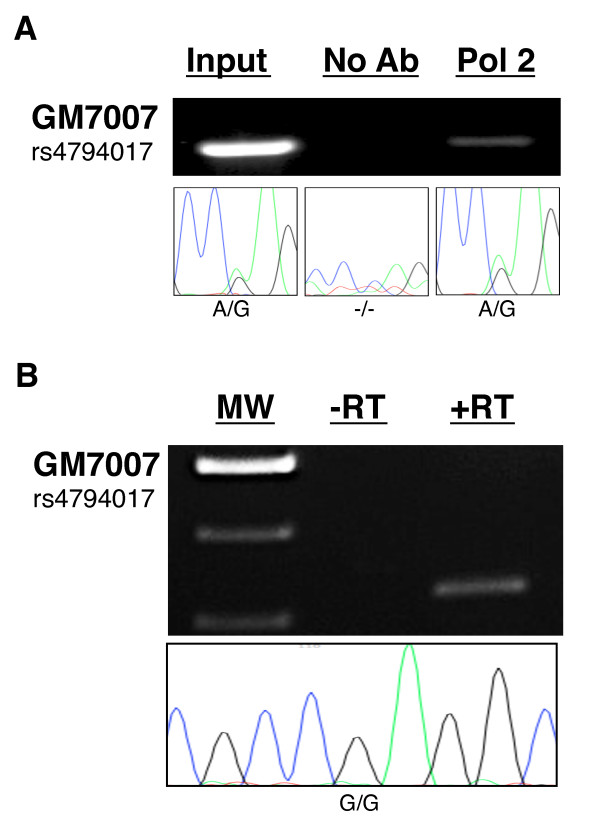
Results: Using array-based chromatin immunoprecipitation, we identified 293 genomic loci that are associated with both CTCF and histone H3 trimethylated at lysine 9 (H3K9me3). A comparison of their genomic positions with those of previously published monoallelically expressed genes revealed no significant overlap between allele-specifically expressed genes and colocalized CTCF/H3K9me3. To analyze the contributions of CTCF and H3K9me3 to gene regulation in more detail, we focused on the monoallelically expressed IGF2BP1 gene. In vitro binding assays using the CTCF target motif at the IGF2BP1 gene, as well as allele-specific analysis of cytosine methylation and CTCF binding, revealed that CTCF does not regulate mono- or biallelic IGF2BP1 expression. Surprisingly, we found that RNA polymerase II is detected on both the maternal and paternal alleles in B lymphoblasts that express IGF2BP1 primarily from one allele. Thus, allele-specific control of RNA polymerase II elongation regulates the allelic bias of IGF2BP1 gene expression.
Conclusions: Colocalization of CTCF and H3K9me3 does not represent a reliable chromatin signature indicative of monoallelic expression. Moreover, association of individual alleles with both active (H3K4me3) and silent (H3K27me3) chromatin modifications (allelic bivalent chromatin) or with RNA polymerase II also fails to identify monoallelically expressed gene loci. The selection of individual alleles for expression occurs in part during transcription elongation.
Figures
References
-
- Bix M, Locksley RM. Independent and epigenetic regulation of the interleukin-4 alleles in CD4+ T cells. Science. 1998;281:1352–1354. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
