

The roles of maternal Vangl2 and aPKC in Xenopus oocyte and embryo patterning
- PMID: 21813572
- PMCID: PMC3160094
- DOI: 10.1242/dev.068866
The roles of maternal Vangl2 and aPKC in Xenopus oocyte and embryo patterning
Abstract
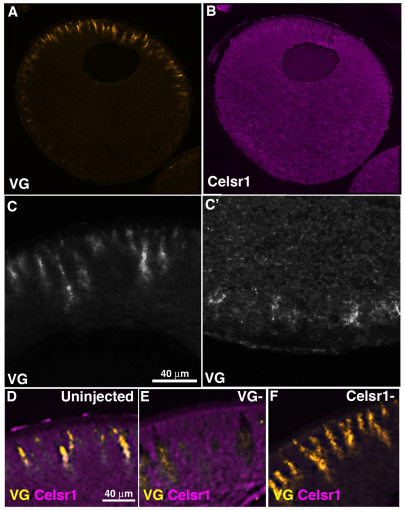
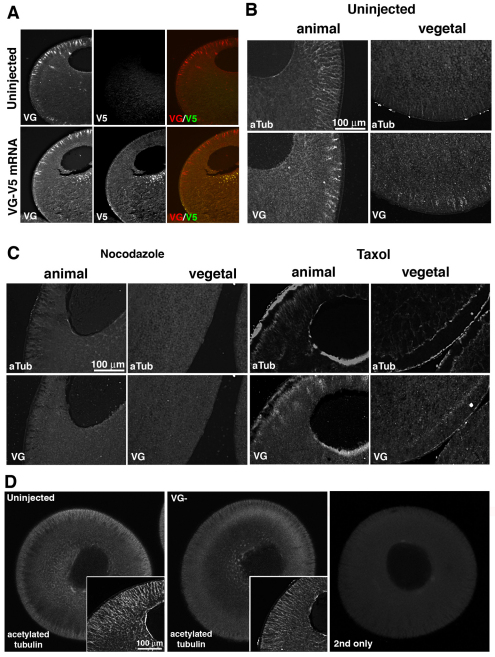
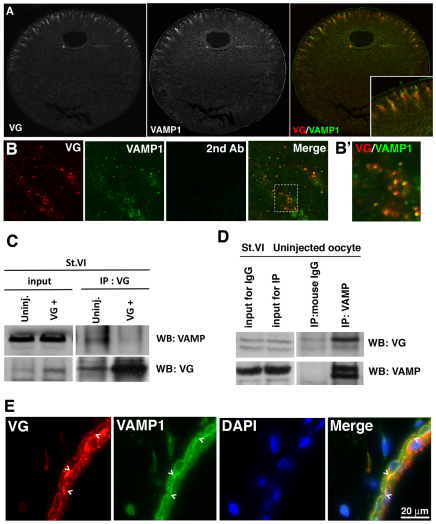
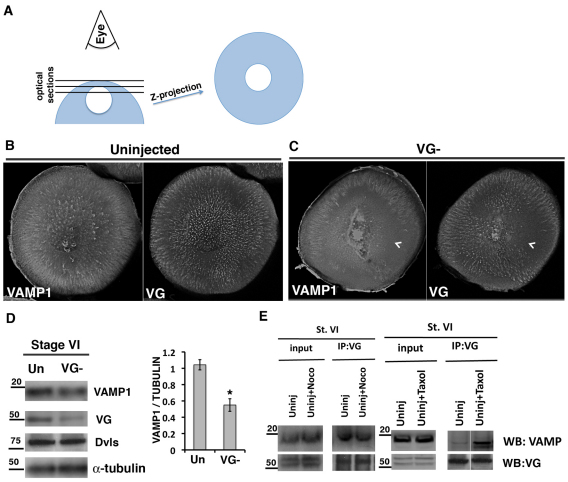
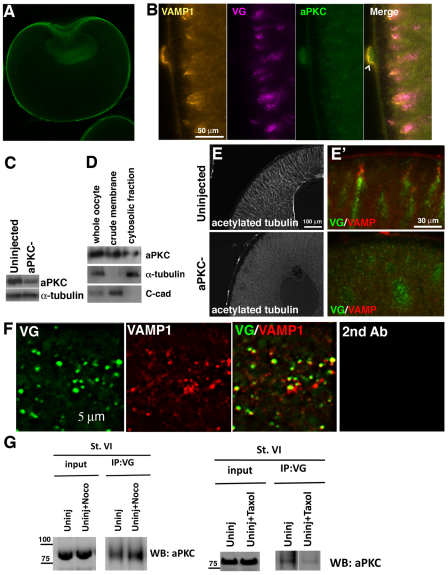
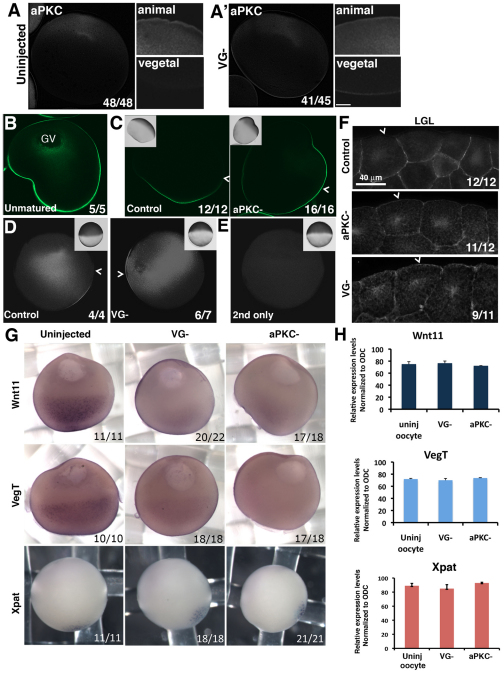
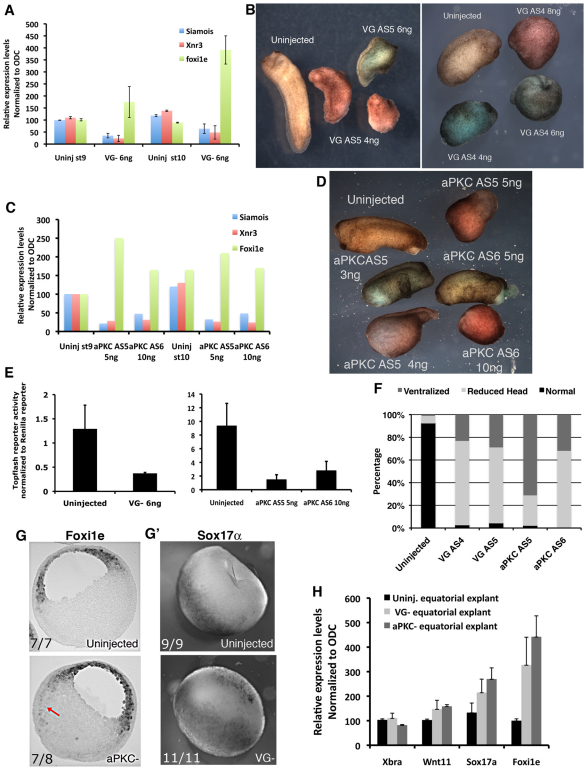
The Xenopus oocyte contains components of both the planar cell polarity and apical-basal polarity pathways, but their roles are not known. Here, we examine the distribution, interactions and functions of the maternal planar cell polarity core protein Vangl2 and the apical-basal complex component aPKC. We show that Vangl2 is distributed in animally enriched islands in the subcortical cytoplasm in full-grown oocytes, where it interacts with a post-Golgi v-SNARE protein, VAMP1, and acetylated microtubules. We find that Vangl2 is required for the stability of VAMP1 as well as for the maintenance of the stable microtubule architecture of the oocyte. We show that Vangl2 interacts with atypical PKC, and that both the acetylated microtubule cytoskeleton and the Vangl2-VAMP1 distribution are dependent on the presence of aPKC. We also demonstrate that aPKC and Vangl2 are required for the cell membrane asymmetry that is established during oocyte maturation, and for the asymmetrical distribution of maternal transcripts for the germ layer and dorsal/ventral determinants VegT and Wnt11. This study demonstrates the interaction and interdependence of Vangl2, VAMP1, aPKC and the stable microtubule cytoskeleton in the oocyte, shows that maternal Vangl2 and aPKC are required for specific oocyte asymmetries and vertebrate embryonic patterning, and points to the usefulness of the oocyte as a model to study the polarity problem.
Figures
References
-
- Aleu J., Blasi J., Solsona C., Marsal J. (2002). Calcium-dependent acetylcholine release from Xenopus oocytes: simultaneous ionic currents and acetylcholine release recordings. Eur. J. Neurosci. 16, 1442-1448 - PubMed
-
- Birsoy B., Kofron M., Schaible K., Wylie C., Heasman J. (2006). Vg 1 is an essential signaling molecule in Xenopus development. Development 133, 15-20 - PubMed
-
- Breckenridge R. A., Mohun T. J., Amaya E. (2001). A role for BMP signalling in heart looping morphogenesis in Xenopus. Dev. Biol. 232, 191-203 - PubMed
-
- Cha S. W., Heasman J. (2010). Using oocytes for Wnt signaling assays: paracrine assays and Wnt-conditioned medium. Methods 51, 52-55 - PubMed
-
- Cha S. W., Tadjuidje E., Tao Q., Wylie C., Heasman J. (2008). Wnt5a and Wnt11 interact in a maternal Dkk1-regulated fashion to activate both canonical and non-canonical signaling in Xenopus axis formation. Development 135, 3719-3729 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
