

Loss of protease dimerization inhibition activity of darunavir is associated with the acquisition of resistance to darunavir by HIV-1
- PMID: 21813613
- PMCID: PMC3196396
- DOI: 10.1128/JVI.05121-11
Loss of protease dimerization inhibition activity of darunavir is associated with the acquisition of resistance to darunavir by HIV-1
Abstract
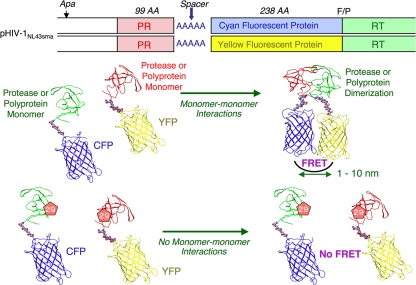
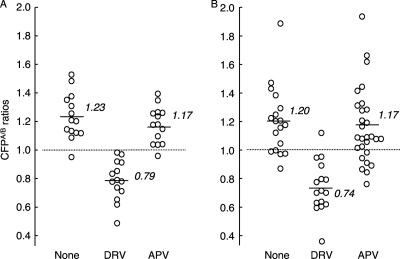
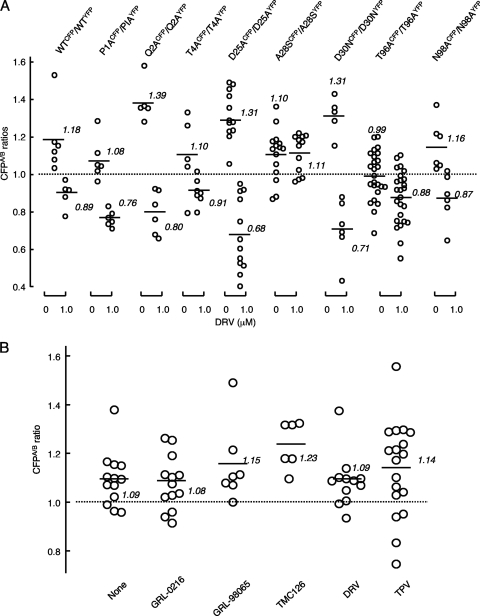
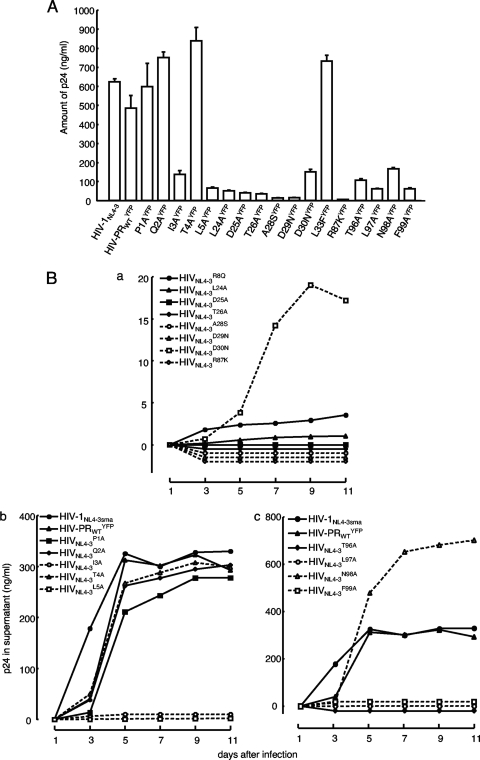
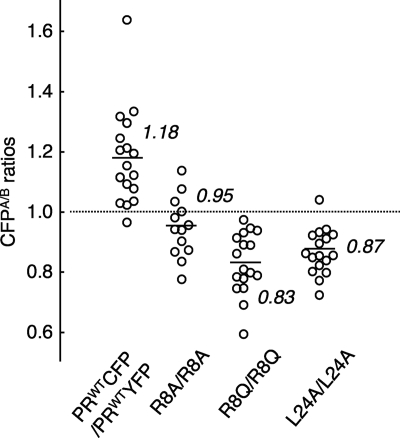
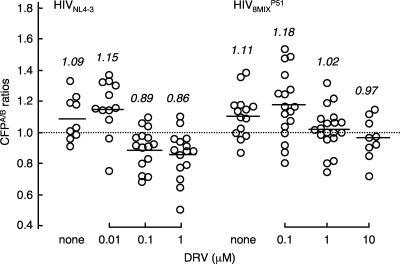
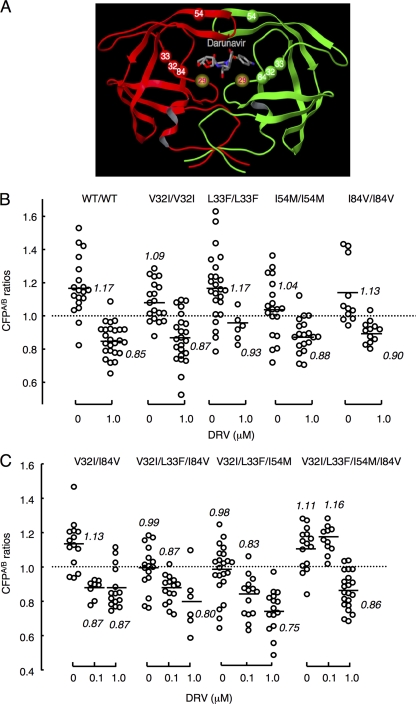
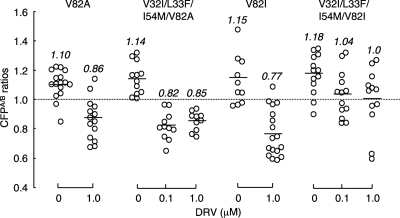
Dimerization of HIV protease is essential for the acquisition of protease's proteolytic activity. We previously identified a group of HIV protease dimerization inhibitors, including darunavir (DRV). In the present work, we examine whether loss of DRV's protease dimerization inhibition activity is associated with HIV development of DRV resistance. Single amino acid substitutions, including I3A, L5A, R8A/Q, L24A, T26A, D29N, R87K, T96A, L97A, and F99A, disrupted protease dimerization, as examined using an intermolecular fluorescence resonance energy transfer (FRET)-based HIV expression assay. All recombinant HIV(NL4-3)-based clones with such a protease dimerization-disrupting substitution failed to replicate. A highly DRV-resistant in vitro-selected HIV variant and clinical HIV strains isolated from AIDS patients failing to respond to DRV-containing antiviral regimens typically had the V32I, L33F, I54M, and I84V substitutions in common in protease. None of up to 3 of the 4 substitutions affected DRV's protease dimerization inhibition, which was significantly compromised by the four combined substitutions. Recombinant infectious clones containing up to 3 of the 4 substitutions remained sensitive to DRV, while a clonal HIV variant with all 4 substitutions proved highly resistant to DRV with a 205-fold 50% effective concentration (EC(50)) difference compared to HIV(NL4-3). The present data suggest that the loss of DRV activity to inhibit protease dimerization represents a novel mechanism contributing to HIV resistance to DRV. The finding that 4 substitutions in PR are required for significant loss of DRV's protease dimerization inhibition should at least partially explain the reason DRV has a high genetic barrier against HIV's acquisition of DRV resistance.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
