

Extracellular signal-regulated kinase signaling in the ventral tegmental area mediates cocaine-induced synaptic plasticity and rewarding effects
- PMID: 21813685
- PMCID: PMC3153072
- DOI: 10.1523/JNEUROSCI.1040-11.2011
Extracellular signal-regulated kinase signaling in the ventral tegmental area mediates cocaine-induced synaptic plasticity and rewarding effects
Abstract
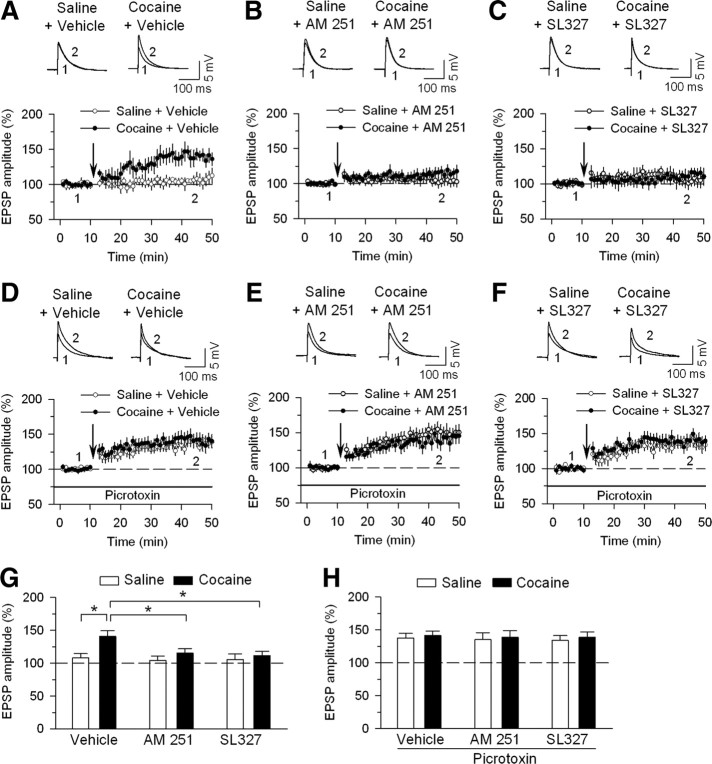
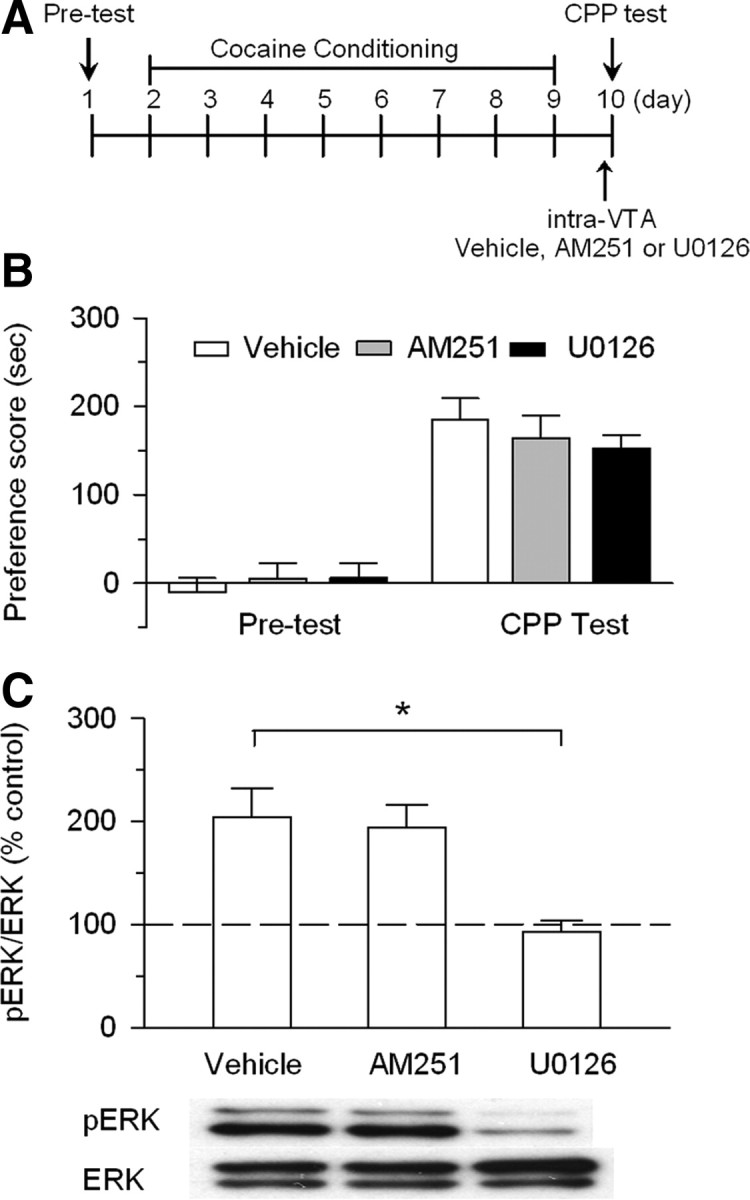
Drugs of abuse such as cocaine induce long-term synaptic plasticity in the reward circuitry, which underlies the formation of drug-associated memories and addictive behavior. We reported previously that repeated cocaine exposure in vivo facilitates long-term potentiation (LTP) in dopamine neurons of the ventral tegmental area (VTA) by reducing the strength of GABAergic inhibition and that endocannabinoid-dependent long-term depression at inhibitory synapses (I-LTD) constitutes a mechanism for cocaine-induced reduction of GABAergic inhibition. The present study investigated the downstream signaling mechanisms and functional consequences of I-LTD in the VTA in the rat. Extracellular signal-regulated kinase (ERK) signaling has been implicated in long-term synaptic plasticity, associative learning, and drug addiction. We tested the hypothesis that VTA ERK activity is required for I-LTD and cocaine-induced long-term synaptic plasticity and behavioral effects. We show that the activation of receptors required for I-LTD increased ERK1/2 phosphorylation and inhibitors of ERK activation blocked I-LTD. We further demonstrate that ERK mediates cocaine-induced reduction of GABAergic inhibition and facilitation of LTP induction. Finally, we show that cocaine conditioned place preference (CPP) training (15 mg/kg; four pairings) increased ERK1/2 phosphorylation in the VTA, while bilateral intra-VTA injections of a CB(1) antagonist or an inhibitor of ERK activation attenuated ERK1/2 phosphorylation and the acquisition, but not the expression, of CPP to cocaine. Our study has identified the CB(1) and ERK signaling cascade as a key mediator of several forms of cocaine-induced synaptic plasticity and provided evidence linking long-term synaptic plasticity in the VTA to rewarding effects of cocaine.
Figures





Similar articles
-
Phosphodiesterase 4 inhibition impairs cocaine-induced inhibitory synaptic plasticity and conditioned place preference.Neuropsychopharmacology. 2012 Oct;37(11):2377-87. doi: 10.1038/npp.2012.93. Epub 2012 Jun 20. Neuropsychopharmacology. 2012. PMID: 22713909 Free PMC article.
-
D2 dopamine receptor activation facilitates endocannabinoid-mediated long-term synaptic depression of GABAergic synaptic transmission in midbrain dopamine neurons via cAMP-protein kinase A signaling.J Neurosci. 2008 Dec 24;28(52):14018-30. doi: 10.1523/JNEUROSCI.4035-08.2008. J Neurosci. 2008. PMID: 19109485 Free PMC article.
-
CaMKII activity in the ventral tegmental area gates cocaine-induced synaptic plasticity in the nucleus accumbens.Neuropsychopharmacology. 2014 Mar;39(4):989-99. doi: 10.1038/npp.2013.299. Epub 2013 Oct 24. Neuropsychopharmacology. 2014. PMID: 24154664 Free PMC article.
-
Synaptic and intrinsic plasticity in the ventral tegmental area after chronic cocaine.Curr Opin Neurobiol. 2019 Feb;54:66-72. doi: 10.1016/j.conb.2018.08.013. Epub 2018 Sep 17. Curr Opin Neurobiol. 2019. PMID: 30237117 Free PMC article. Review.
-
LTP of GABAergic synapses in the ventral tegmental area and beyond.J Physiol. 2008 Mar 15;586(6):1487-93. doi: 10.1113/jphysiol.2007.148098. Epub 2007 Dec 13. J Physiol. 2008. PMID: 18079157 Free PMC article. Review.
Cited by
-
BDNF interacts with endocannabinoids to regulate cocaine-induced synaptic plasticity in mouse midbrain dopamine neurons.J Neurosci. 2015 Mar 11;35(10):4469-81. doi: 10.1523/JNEUROSCI.2924-14.2015. J Neurosci. 2015. PMID: 25762688 Free PMC article.
-
Inactivation of ERK1/2 Signaling in Dopaminergic Neurons by Map Kinase Phosphatase MKP3 Regulates Dopamine Signaling and Motivation for Cocaine.J Neurosci. 2024 Jan 31;44(5):e0727232023. doi: 10.1523/JNEUROSCI.0727-23.2023. J Neurosci. 2024. PMID: 38296649 Free PMC article.
-
Stress as a Risk Factor for Substance Use Disorders: A Mini-Review of Molecular Mediators.Front Behav Neurosci. 2018 Dec 21;12:309. doi: 10.3389/fnbeh.2018.00309. eCollection 2018. Front Behav Neurosci. 2018. PMID: 30622460 Free PMC article. Review.
-
The extracellular signal-regulated kinase 1/2 pathway in neurological diseases: A potential therapeutic target (Review).Int J Mol Med. 2017 Jun;39(6):1338-1346. doi: 10.3892/ijmm.2017.2962. Epub 2017 Apr 21. Int J Mol Med. 2017. PMID: 28440493 Free PMC article. Review.
-
Allosteric Modulation: An Alternate Approach Targeting the Cannabinoid CB1 Receptor.Med Res Rev. 2017 May;37(3):441-474. doi: 10.1002/med.21418. Epub 2016 Nov 23. Med Res Rev. 2017. PMID: 27879006 Free PMC article. Review.
References
-
- Atkins CM, Selcher JC, Petraitis JJ, Trzaskos JM, Sweatt JD. The MAPK cascade is required for mammalian associative learning. Nat Neurosci. 1998;1:602–609. - PubMed
-
- Bolshakov VY, Carboni L, Cobb MH, Siegelbaum SA, Belardetti F. Dual MAP kinase pathways mediate opposing forms of long-term plasticity at CA3-CA1 synapses. Nat Neurosci. 2000;3:1107–1112. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous