

The roles of conserved and nonconserved cysteinyl residues in the oligomerization and function of mammalian prestin
- PMID: 21813750
- PMCID: PMC3214107
- DOI: 10.1152/jn.00496.2011
The roles of conserved and nonconserved cysteinyl residues in the oligomerization and function of mammalian prestin
Abstract
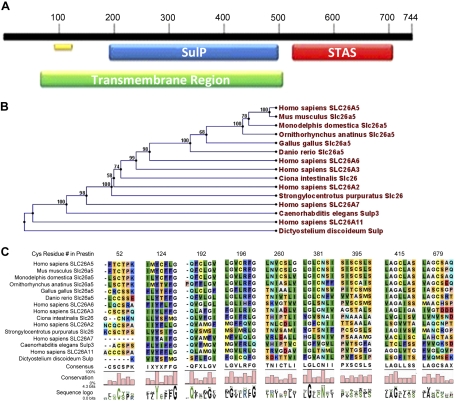
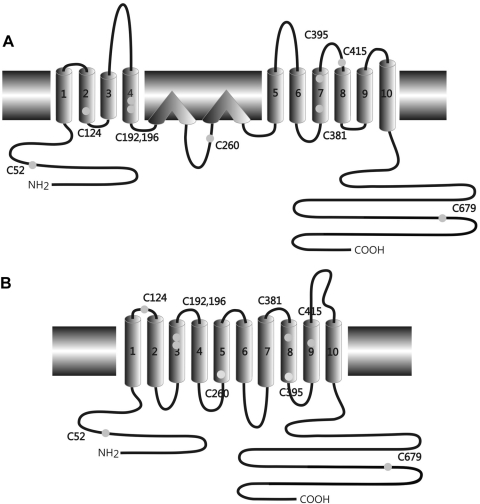
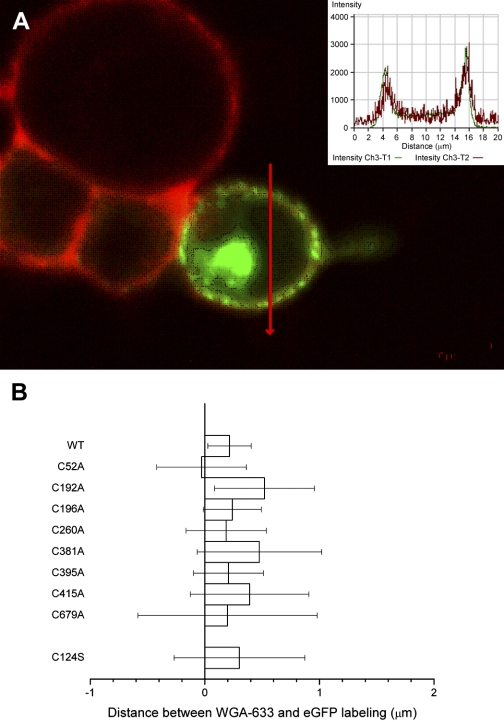
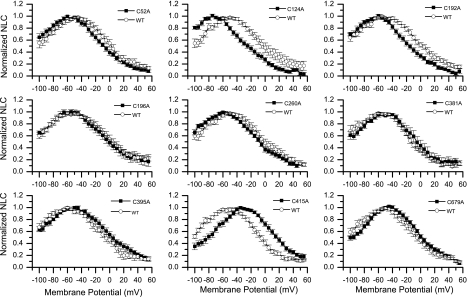
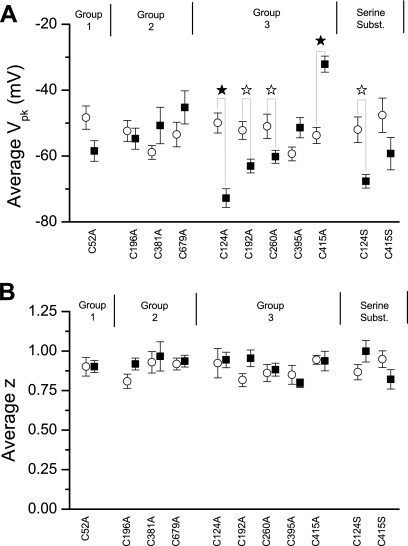
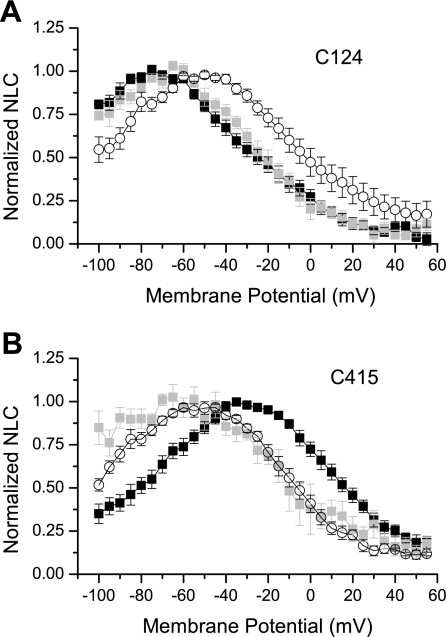
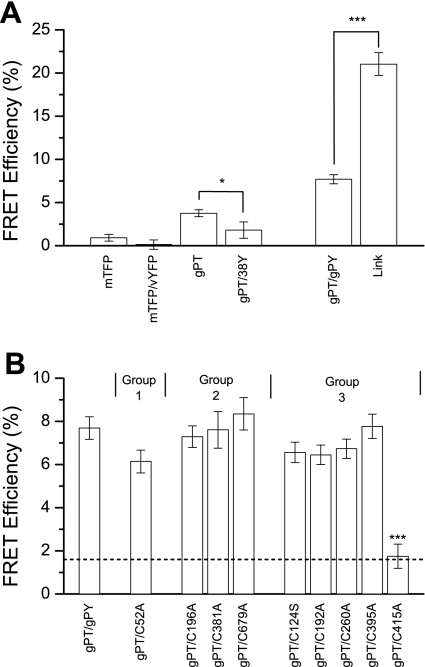
The creation of several prestin knockout and knockin mouse lines has demonstrated the importance of the intrinsic outer hair cell membrane protein prestin to mammalian hearing. However, the structure of prestin remains largely unknown, with even its major features in dispute. Several studies have suggested that prestin forms homo-oligomers that may be stabilized by disulfide bonds. Our phylogenetic analysis of prestin sequences across chordate classes suggested that the cysteinyl residues could be divided into three groups, depending on the extent of their conservation between prestin orthologs and paralogs or homologs. An alanine scan functional analysis was performed of all nine cysteinyl positions in mammalian prestin. Prestin function was assayed by measurement of prestin-associated nonlinear capacitance. Of the nine cysteine-alanine substitution mutations, all were properly membrane targeted and all demonstrated nonlinear capacitance. Four mutations (C124A, C192A, C260A, and C415A), all in nonconserved cysteinyl residues, significantly differed in their nonlinear capacitance properties compared with wild-type prestin. In the two most severely disrupted mutations, substitution of the polar residue seryl for cysteinyl restored normal function in one (C415S) but not the other (C124S). We assessed the relationship of prestin oligomerization to cysteine position using fluorescence resonance energy transfer. With one exception, cysteine-alanine substitutions did not significantly alter prestin-prestin interactions. The exception was C415A, one of the two nonconserved cysteinyl residues whose mutation to alanine caused the most disruption in function. We suggest that no disulfide bond is essential for prestin function. However, C415 likely participates by hydrogen bonding in both nonlinear capacitance and oligomerization.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources