

Antagonism of LIN-17/Frizzled and LIN-18/Ryk in nematode vulva induction reveals evolutionary alterations in core developmental pathways
- PMID: 21814488
- PMCID: PMC3144188
- DOI: 10.1371/journal.pbio.1001110
Antagonism of LIN-17/Frizzled and LIN-18/Ryk in nematode vulva induction reveals evolutionary alterations in core developmental pathways
Abstract
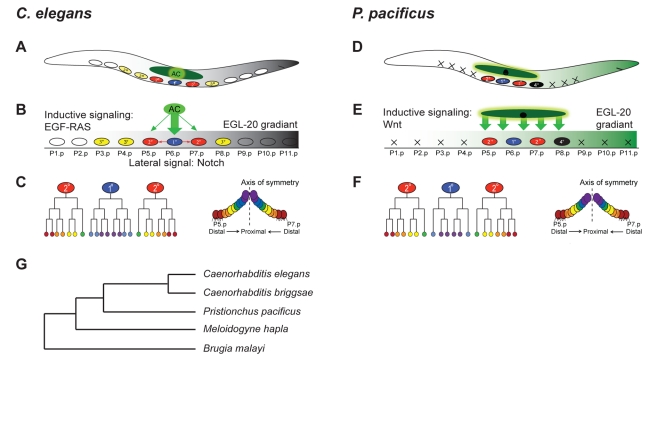
Most diversity in animals and plants results from the modification of already existing structures. Many organ systems, for example, are permanently modified during evolution to create developmental and morphological diversity, but little is known about the evolution of the underlying developmental mechanisms. The theory of developmental systems drift proposes that the development of conserved morphological structures can involve large-scale modifications in their regulatory mechanisms. We test this hypothesis by comparing vulva induction in two genetically tractable nematodes, Caenorhabditis elegans and Pristionchus pacificus. Previous work indicated that the vulva is induced by epidermal growth factor (EGF)/RAS and WNT signaling in Caenorhabditis and Pristionchus, respectively. Here, we show that the evolution of vulva induction involves major molecular alterations and that this shift of signaling pathways involves a novel wiring of WNT signaling and the acquisition of novel domains in otherwise conserved receptors in Pristionchus vulva induction. First, Ppa-LIN-17/Frizzled acts as an antagonist of WNT signaling and suppresses the ligand Ppa-EGL-20 by ligand sequestration. Second, Ppa-LIN-18/Ryk transmits WNT signaling and requires inhibitory SH3 domain binding motifs, unknown from Cel-LIN-18/Ryk. Third, Ppa-LIN-18/Ryk signaling involves Axin and β-catenin and Ppa-axl-1/Axin is epistatic to Ppa-lin-18/Ryk. These results confirm developmental system drift as an important theory for the evolution of organ systems and they highlight the significance of protein modularity in signal transduction and the dynamics of signaling networks.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures






Comment in
-
Different paths, same structure: "developmental systems drift" at work.PLoS Biol. 2011 Jul;9(7):e1001113. doi: 10.1371/journal.pbio.1001113. Epub 2011 Jul 26. PLoS Biol. 2011. PMID: 21814491 Free PMC article. No abstract available.
-
Evo-devo: Hidden rewiring comes to light.Nat Rev Genet. 2011 Aug 9;12(9):586. doi: 10.1038/nrg3060. Nat Rev Genet. 2011. PMID: 21826064 No abstract available.
Similar articles
-
Wnt signaling induces vulva development in the nematode Pristionchus pacificus.Curr Biol. 2008 Jan 22;18(2):142-6. doi: 10.1016/j.cub.2007.12.048. Curr Biol. 2008. PMID: 18207741
-
The pax-3 gene is involved in vulva formation in Pristionchus pacificus and is a target of the Hox gene lin-39.Development. 2007 Sep;134(17):3111-9. doi: 10.1242/dev.008375. Epub 2007 Jul 25. Development. 2007. PMID: 17652349
-
Evolution of regulatory networks: nematode vulva induction as an example of developmental systems drift.Adv Exp Med Biol. 2012;751:79-91. doi: 10.1007/978-1-4614-3567-9_4. Adv Exp Med Biol. 2012. PMID: 22821454
-
Wnt signaling went derailed again: a new track via the LIN-18 receptor?Cell. 2004 Sep 17;118(6):668-70. doi: 10.1016/j.cell.2004.09.009. Cell. 2004. PMID: 15369666 Review.
-
Vulval development.WormBook. 2005 Jun 25:1-28. doi: 10.1895/wormbook.1.6.1. WormBook. 2005. PMID: 18050418 Free PMC article. Review.
Cited by
-
Evolution of New cis-Regulatory Motifs Required for Cell-Specific Gene Expression in Caenorhabditis.PLoS Genet. 2016 Sep 2;12(9):e1006278. doi: 10.1371/journal.pgen.1006278. eCollection 2016 Sep. PLoS Genet. 2016. PMID: 27588814 Free PMC article.
-
Characterization of the Pristionchus pacificus "epigenetic toolkit" reveals the evolutionary loss of the histone methyltransferase complex PRC2.Genetics. 2024 May 7;227(1):iyae041. doi: 10.1093/genetics/iyae041. Genetics. 2024. PMID: 38513719 Free PMC article.
-
Cis-regulatory programs in the development and evolution of vertebrate paired appendages.Semin Cell Dev Biol. 2016 Sep;57:31-39. doi: 10.1016/j.semcdb.2016.01.015. Epub 2016 Jan 16. Semin Cell Dev Biol. 2016. PMID: 26783722 Free PMC article. Review.
-
Genome-wide analysis of germline signaling genes regulating longevity and innate immunity in the nematode Pristionchus pacificus.PLoS Pathog. 2012;8(8):e1002864. doi: 10.1371/journal.ppat.1002864. Epub 2012 Aug 9. PLoS Pathog. 2012. PMID: 22912581 Free PMC article.
-
Cryptic variation in vulva development by cis-regulatory evolution of a HAIRY-binding site.Nat Commun. 2013;4:1714. doi: 10.1038/ncomms2711. Nat Commun. 2013. PMID: 23591881
References
-
- Raff R. A. The shape of life: genes, development, and the evolution of animal form. Chicago: The University of Chicago Press; 1996.
-
- Bento G, Ogawa A, Sommer R. J. Co-option of the hormone-signaling module dafachronic acid-DAF-12 in nematode evolution. Nature. 2010;466:494–497. - PubMed
-
- Schmidt-Rhaesa A. The evolution of organ systems. New York: Oxford University Press; 2007.
-
- True J. R, Haag E. S. Developmental system drift and flexibility in evolutionary trajectories. Evol Dev. 2001;3:109–119. - PubMed
-
- Sommer R. J. The future of evo-devo: model systems and evolutionary theory. Nat Rev Genet. 2009;10:416–422. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
