

The Eps8/IRSp53/VASP network differentially controls actin capping and bundling in filopodia formation
- PMID: 21814501
- PMCID: PMC3140970
- DOI: 10.1371/journal.pcbi.1002088
The Eps8/IRSp53/VASP network differentially controls actin capping and bundling in filopodia formation
Abstract
There is a body of literature that describes the geometry and the physics of filopodia using either stochastic models or partial differential equations and elasticity and coarse-grained theory. Comparatively, there is a paucity of models focusing on the regulation of the network of proteins that control the formation of different actin structures. Using a combination of in-vivo and in-vitro experiments together with a system of ordinary differential equations, we focused on a small number of well-characterized, interacting molecules involved in actin-dependent filopodia formation: the actin remodeler Eps8, whose capping and bundling activities are a function of its ligands, Abi-1 and IRSp53, respectively; VASP and Capping Protein (CP), which exert antagonistic functions in controlling filament elongation. The model emphasizes the essential role of complexes that contain the membrane deforming protein IRSp53, in the process of filopodia initiation. This model accurately accounted for all observations, including a seemingly paradoxical result whereby genetic removal of Eps8 reduced filopodia in HeLa, but increased them in hippocampal neurons, and generated quantitative predictions, which were experimentally verified. The model further permitted us to explain how filopodia are generated in different cellular contexts, depending on the dynamic interaction established by Eps8, IRSp53 and VASP with actin filaments, thus revealing an unexpected plasticity of the signaling network that governs the multifunctional activities of its components in the formation of filopodia.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

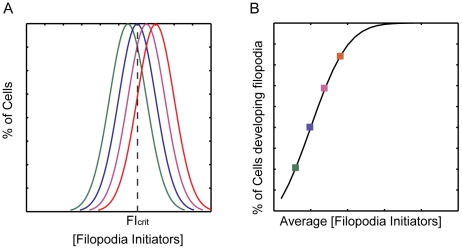
 . The concentration of FI required for initiating the positive feedback loop (FIcrit) is shown as a dotted line. As μ increases (different colored curves) the fraction of cells with FI>FIcrit increases. b. Fraction of cells in a population with FI>FIcrit as a function of the average FI concentration μ. Different color squares represent the fraction of cells for the different Gaussians shown in A. To calculate the amount of cells with FI>FIcrit, we simply integrate the Gaussian from FIcrit to infinite,
. The concentration of FI required for initiating the positive feedback loop (FIcrit) is shown as a dotted line. As μ increases (different colored curves) the fraction of cells with FI>FIcrit increases. b. Fraction of cells in a population with FI>FIcrit as a function of the average FI concentration μ. Different color squares represent the fraction of cells for the different Gaussians shown in A. To calculate the amount of cells with FI>FIcrit, we simply integrate the Gaussian from FIcrit to infinite,  .
.

References
-
- Mattila PK, Lappalainen P. Filopodia: molecular architecture and cellular functions. Nat Rev Mol Cell Biol. 2008;9:446–454. - PubMed
-
- Lebrand C, Dent EW, Strasser GA, Lanier LM, Krause M, et al. Critical role of Ena/VASP proteins for filopodia formation in neurons and in function downstream of netrin-1. Neuron. 2004;42:37–49. - PubMed
