

The Min oscillator uses MinD-dependent conformational changes in MinE to spatially regulate cytokinesis
- PMID: 21816275
- PMCID: PMC3155264
- DOI: 10.1016/j.cell.2011.06.042
The Min oscillator uses MinD-dependent conformational changes in MinE to spatially regulate cytokinesis
Abstract
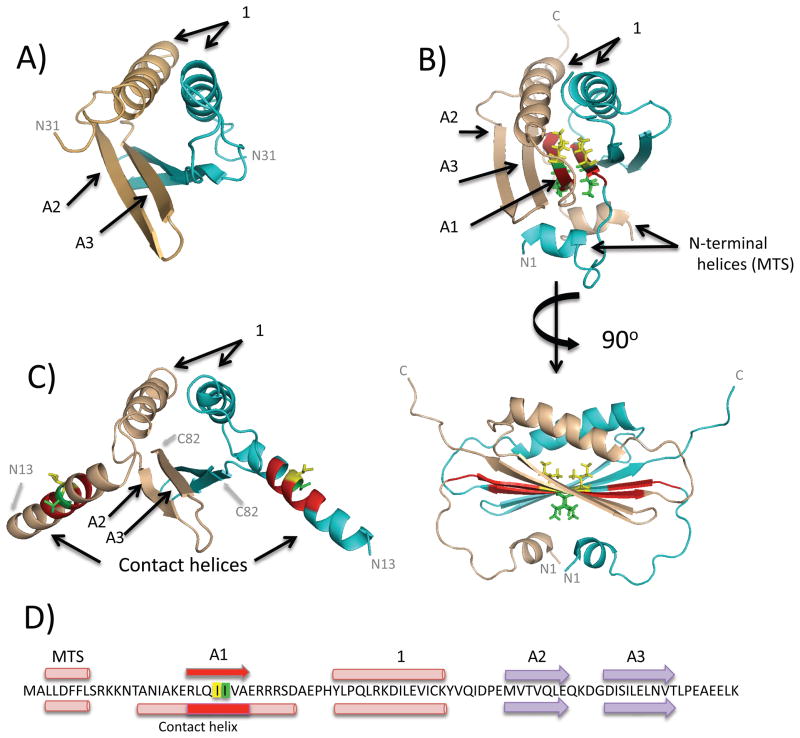
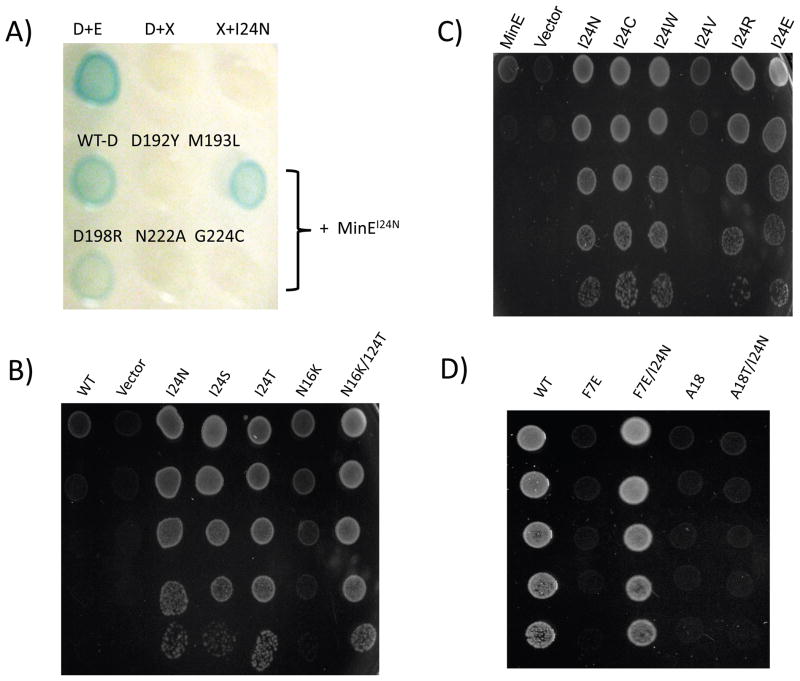
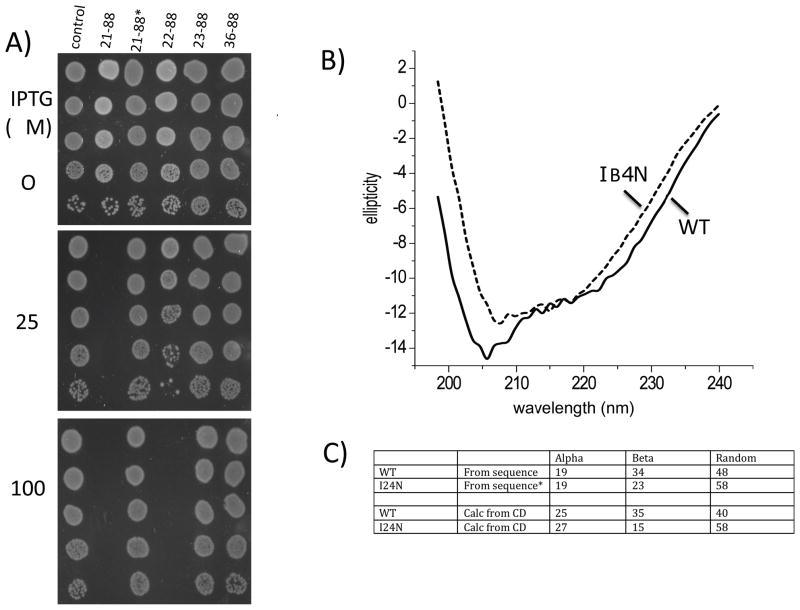
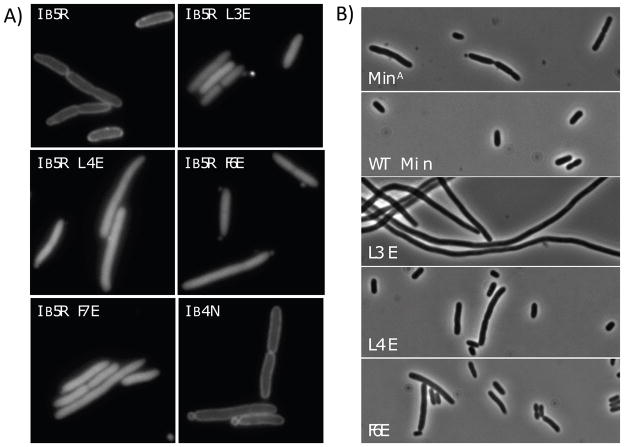
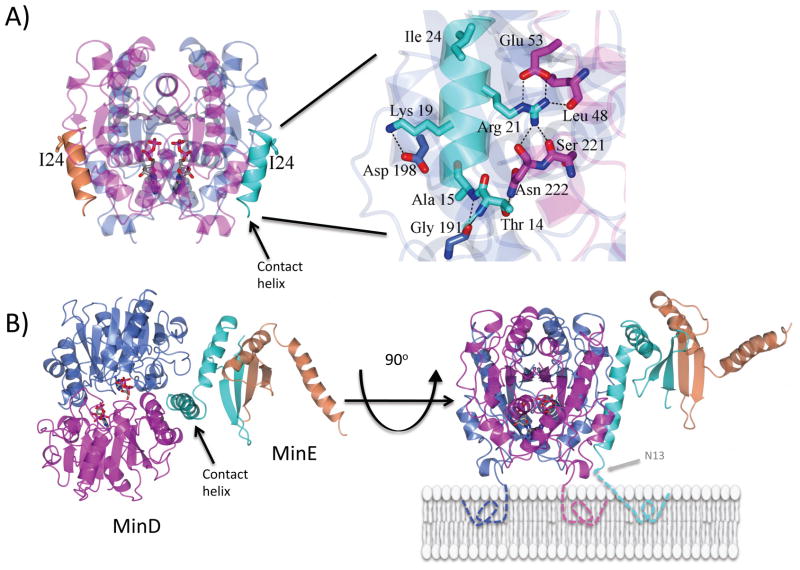
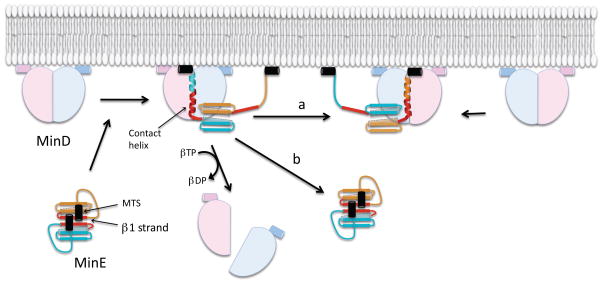
In E. coli, MinD recruits MinE to the membrane, leading to a coupled oscillation required for spatial regulation of the cytokinetic Z ring. How these proteins interact, however, is not clear because the MinD-binding regions of MinE are sequestered within a six-stranded β sheet and masked by N-terminal helices. minE mutations that restore interaction between some MinD and MinE mutants were isolated. These mutations alter the MinE structure leading to release of the MinD-binding regions and the N-terminal helices that bind the membrane. Crystallization of MinD-MinE complexes revealed a four-stranded β sheet MinE dimer with the released β strands (MinD-binding regions) converted to α helices bound to MinD dimers. These results identify the MinD-dependent conformational changes in MinE that convert it from a latent to an active form and lead to a model of how MinE persists at the MinD-membrane surface.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
References
-
- Andrade MA, Chacon P, Merelo JJ, Moran F. Evaluation of secondary structure of proteins from UV circular dichroism spectra using an unsupervised learning neural network. Protein Eng. 1993;6:383–390. - PubMed
-
- de Boer PA, Crossley RE, Rothfield LI. A division inhibitor and a topological specificity factor coded for by the minicell locus determine proper placement of the division septum in E. coli. Cell. 1989;56:641–649. - PubMed
-
- Evans P. Scaling and assessment of data quality. Acta Crystallogr D Biol Crystallogr. 2006;62:72–82. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
