

CXCL16 recruits bone marrow-derived fibroblast precursors in renal fibrosis
- PMID: 21816936
- PMCID: PMC3187185
- DOI: 10.1681/ASN.2010080881
CXCL16 recruits bone marrow-derived fibroblast precursors in renal fibrosis
Abstract
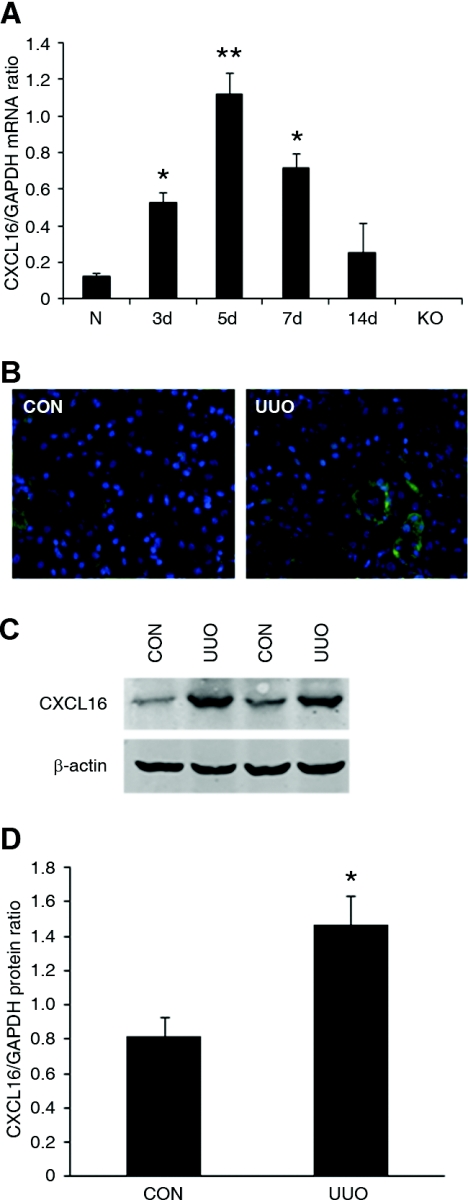
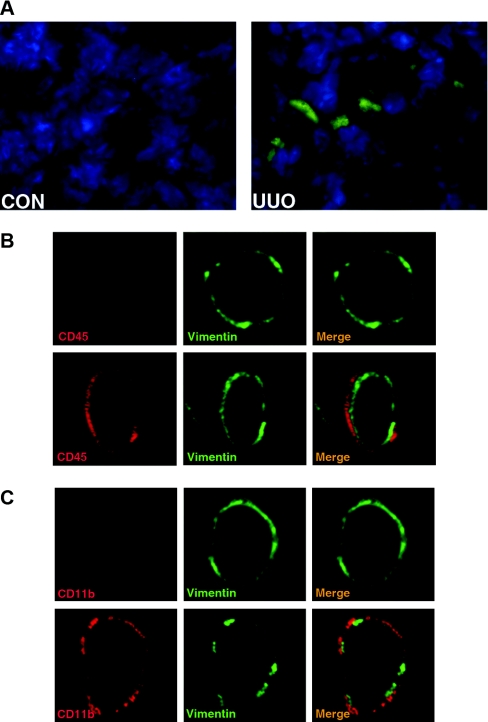
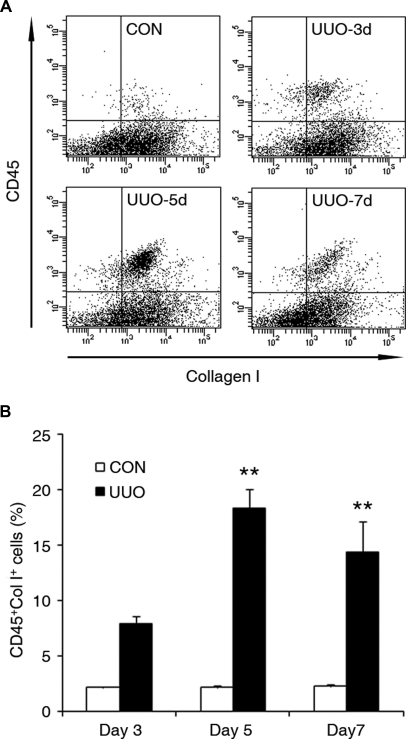
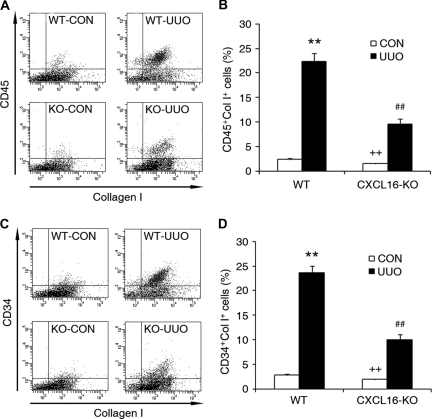
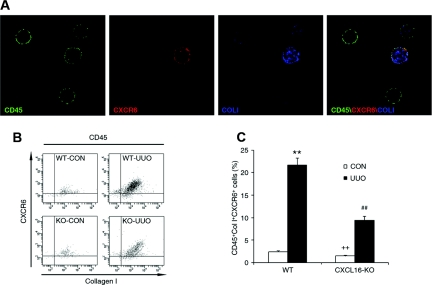
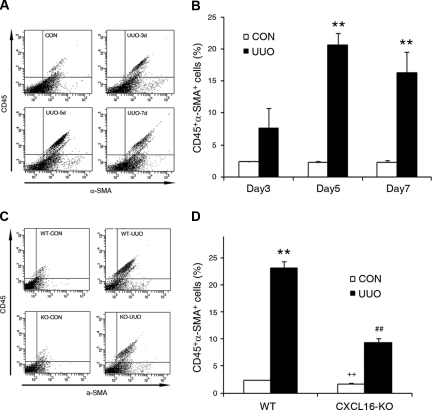
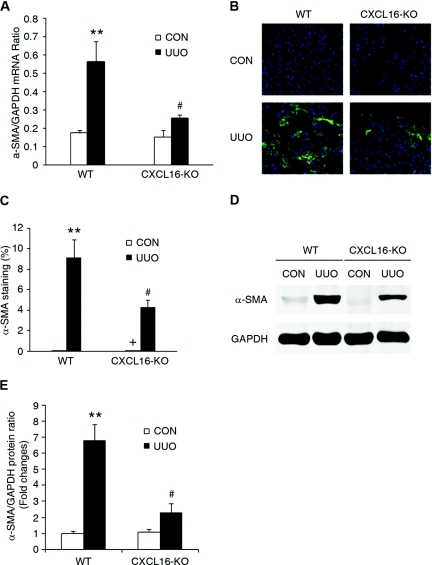
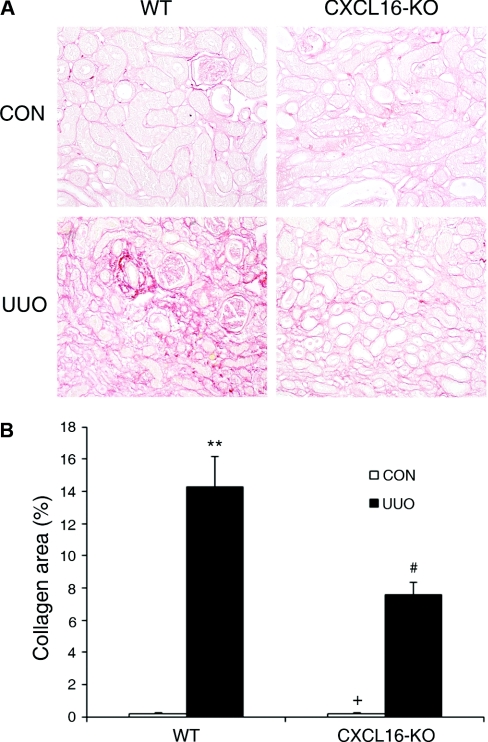
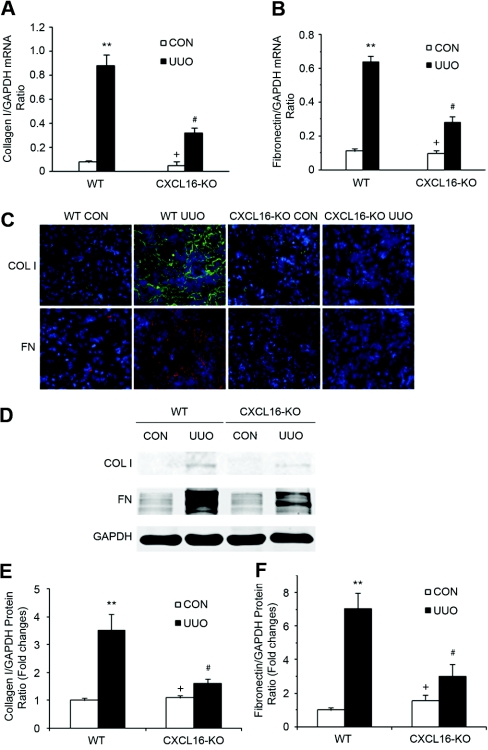
Although fibroblasts are responsible for the production and deposition of extracellular matrix in renal fibrosis, their origin is controversial. Circulating fibroblast precursors may contribute to the pathogenesis of renal fibrosis, but the signaling mechanisms underlying the recruitment of bone marrow-derived fibroblast precursors into the kidney in response to injury are incompletely understood. Here, in the unilateral ureteral obstruction model of renal fibrosis, tubular epithelial cells upregulated the chemokine CXCL16 in obstructed kidneys, and circulating fibroblast precursors expressed the CXCL16 receptor, CXCR6. Compared with wild-type mice, CXCL16-knockout mice accumulated significantly fewer bone marrow-derived fibroblast precursors in obstructed kidneys. CXCL16-knockout mice also exhibited significantly fewer CD45-, collagen I-, and CXCR6-triple-positive fibroblast precursors in injured kidneys. Furthermore, targeted deletion of CXCL16 inhibited myofibroblast activation, reduced collagen deposition, and suppressed expression of collagen I and fibronectin. In conclusion, CXCL16 contributes to the pathogenesis of renal fibrosis by recruiting bone marrow-derived fibroblast precursors.
Figures
References
-
- Schainuck LI, Striker GE, Cutler RE, Benditt EP: Structural-functional correlations in renal disease. II. The correlations. Hum Pathol 1: 631–641, 1970 - PubMed
-
- Nath KA: The tubulointerstitium in progressive renal disease. Kidney Int 54: 992–994, 1998 - PubMed
-
- Eddy AA: Molecular basis of renal fibrosis. Pediatr Nephrol 15: 290–301, 2000 - PubMed
-
- Neilson EG: Mechanisms of disease: Fibroblasts—A new look at an old problem. Nat Clin Pract Nephrol 2: 101–108, 2006 - PubMed
-
- Strutz F, Muller GA: Renal fibrosis and the origin of the renal fibroblast. Nephrol Dial Transplant 21: 3368–3370, 2006 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
