

The Vein Patterning 1 (VEP1) gene family laterally spread through an ecological network
- PMID: 21818306
- PMCID: PMC3144213
- DOI: 10.1371/journal.pone.0022279
The Vein Patterning 1 (VEP1) gene family laterally spread through an ecological network
Abstract
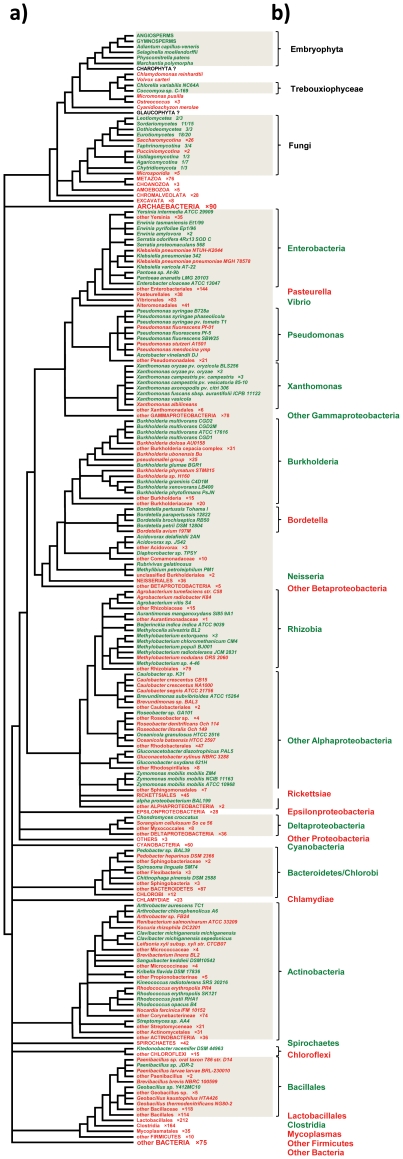
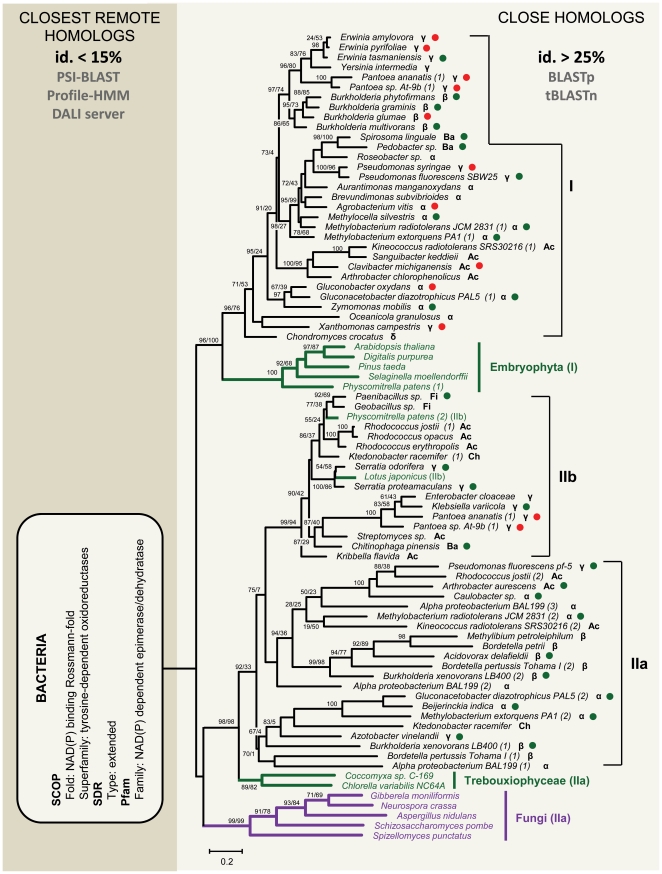
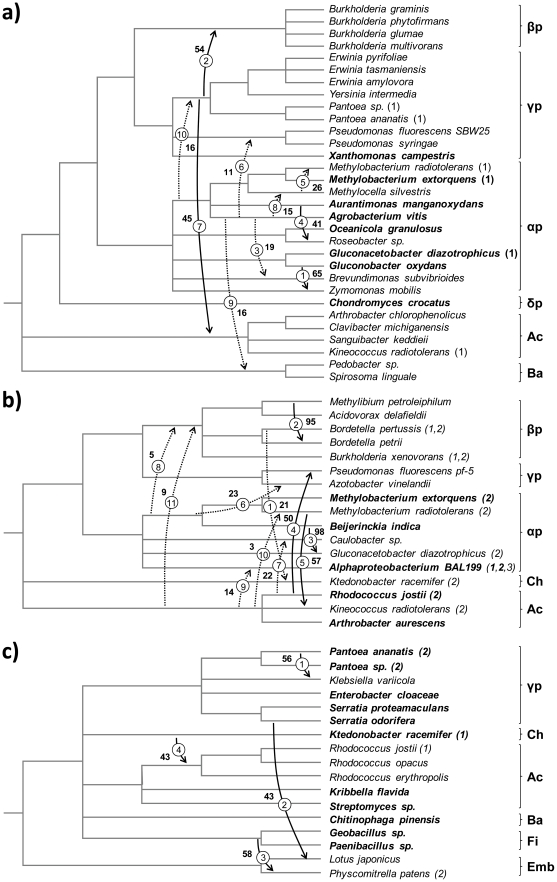
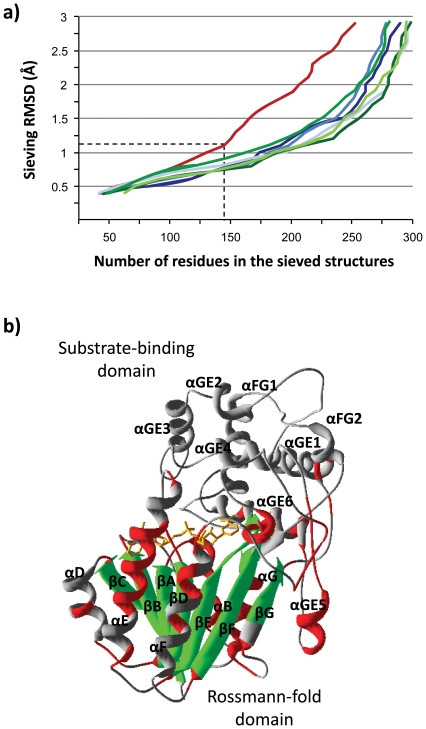
Lateral gene transfer (LGT) is a major evolutionary mechanism in prokaryotes. Knowledge about LGT--particularly, multicellular--eukaryotes has only recently started to accumulate. A widespread assumption sees the gene as the unit of LGT, largely because little is yet known about how LGT chances are affected by structural/functional features at the subgenic level. Here we trace the evolutionary trajectory of VEin Patterning 1, a novel gene family known to be essential for plant development and defense. At the subgenic level VEP1 encodes a dinucleotide-binding Rossmann-fold domain, in common with members of the short-chain dehydrogenase/reductase (SDR) protein family. We found: i) VEP1 likely originated in an aerobic, mesophilic and chemoorganotrophic α-proteobacterium, and was laterally propagated through nets of ecological interactions, including multiple LGTs between phylogenetically distant green plant/fungi-associated bacteria, and five independent LGTs to eukaryotes. Of these latest five transfers, three are ancient LGTs, implicating an ancestral fungus, the last common ancestor of land plants and an ancestral trebouxiophyte green alga, and two are recent LGTs to modern embryophytes. ii) VEP1's rampant LGT behavior was enabled by the robustness and broad utility of the dinucleotide-binding Rossmann-fold, which provided a platform for the evolution of two unprecedented departures from the canonical SDR catalytic triad. iii) The fate of VEP1 in eukaryotes has been different in different lineages, being ubiquitous and highly conserved in land plants, whereas fungi underwent multiple losses. And iv) VEP1-harboring bacteria include non-phytopathogenic and phytopathogenic symbionts which are non-randomly distributed with respect to the type of harbored VEP1 gene. Our findings suggest that VEP1 may have been instrumental for the evolutionary transition of green plants to land, and point to a LGT-mediated 'Trojan Horse' mechanism for the evolution of bacterial pathogenesis against plants. VEP1 may serve as tool for revealing microbial interactions in plant/fungi-associated environments.
Conflict of interest statement
Figures
Similar articles
-
Grafting or pruning in the animal tree: lateral gene transfer and gene loss?BMC Genomics. 2018 Jun 18;19(1):470. doi: 10.1186/s12864-018-4832-5. BMC Genomics. 2018. PMID: 29914363 Free PMC article.
-
Lateral gene transfer between prokaryotes and eukaryotes.Exp Cell Res. 2017 Sep 15;358(2):421-426. doi: 10.1016/j.yexcr.2017.02.009. Epub 2017 Feb 9. Exp Cell Res. 2017. PMID: 28189637 Free PMC article. Review.
-
Old genes in new places: A taxon-rich analysis of interdomain lateral gene transfer events.PLoS Genet. 2022 Jun 22;18(6):e1010239. doi: 10.1371/journal.pgen.1010239. eCollection 2022 Jun. PLoS Genet. 2022. PMID: 35731825 Free PMC article.
-
Paths of lateral gene transfer of lysyl-aminoacyl-tRNA synthetases with a unique evolutionary transition stage of prokaryotes coding for class I and II varieties by the same organisms.BMC Evol Biol. 2006 Mar 12;6:22. doi: 10.1186/1471-2148-6-22. BMC Evol Biol. 2006. PMID: 16529662 Free PMC article.
-
Lateral gene transfers and the origins of the eukaryote proteome: a view from microbial parasites.Curr Opin Microbiol. 2015 Feb;23:155-62. doi: 10.1016/j.mib.2014.11.018. Epub 2014 Dec 5. Curr Opin Microbiol. 2015. PMID: 25483352 Free PMC article. Review.
Cited by
-
RNAi-mediated gene knockdown of progesterone 5β-reductases in Digitalis lanata reduces 5β-cardenolide content.Plant Cell Rep. 2021 Sep;40(9):1631-1646. doi: 10.1007/s00299-021-02707-3. Epub 2021 Jun 19. Plant Cell Rep. 2021. PMID: 34146141 Free PMC article.
-
Ancient horizontal transfer of transaldolase-like protein gene and its role in plant vascular development.New Phytol. 2015 Apr;206(2):807-16. doi: 10.1111/nph.13183. Epub 2014 Nov 24. New Phytol. 2015. PMID: 25420550 Free PMC article.
-
The plant short-chain dehydrogenase (SDR) superfamily: genome-wide inventory and diversification patterns.BMC Plant Biol. 2012 Nov 20;12:219. doi: 10.1186/1471-2229-12-219. BMC Plant Biol. 2012. PMID: 23167570 Free PMC article.
-
Ancestor of land plants acquired the DNA-3-methyladenine glycosylase (MAG) gene from bacteria through horizontal gene transfer.Sci Rep. 2017 Aug 24;7(1):9324. doi: 10.1038/s41598-017-05066-w. Sci Rep. 2017. PMID: 28839126 Free PMC article.
-
Knockout of Arabidopsis thaliana VEP1, Encoding a PRISE (Progesterone 5β-Reductase/Iridoid Synthase-Like Enzyme), Leads to Metabolic Changes in Response to Exogenous Methyl Vinyl Ketone (MVK).Metabolites. 2021 Dec 23;12(1):11. doi: 10.3390/metabo12010011. Metabolites. 2021. PMID: 35050133 Free PMC article.
References
-
- Frost LS, Leplae R, Summers AO, Toussaint A. Mobile genetic elements: the agents of open source evolution. Nat Rev Microbiol. 2005;3:722–732. - PubMed
-
- Thomas CM, Nielsen KM. Mechanisms of, and barriers to, horizontal gene transfer between bacteria. Nat Rev Microbiol. 2005;3:711–721. - PubMed
-
- Lang AS, Beatty JT. Importance of widespread gene transfer agent genes in alphaproteobacteria. Trends Microbiol. 2007;15:54–62. - PubMed
-
- Doolittle WF. Phylogenetic classification and the universal tree. Science. 1999;284:2124–2129. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
