

Review
doi: 10.1007/s00281-011-0286-4.
Epub 2011 Aug 6.
Platelets: versatile effector cells in hemostasis, inflammation, and the immune continuum
Affiliations
- PMID: 21818701
- PMCID: PMC4334392
- DOI: 10.1007/s00281-011-0286-4
Item in Clipboard
Review
Platelets: versatile effector cells in hemostasis, inflammation, and the immune continuum
Semin Immunopathol.
2012 Jan.
Abstract
Platelets are chief effector cells in hemostasis. In addition, however, their specializations include activities and intercellular interactions that make them key effectors in inflammation and in the continuum of innate and adaptive immunity. This review focuses on the immune features of human platelets and platelets from experimental animals and on interactions between inflammatory, immune, and hemostatic activities of these anucleate but complex and versatile cells. The experimental findings and evidence for physiologic immune functions include previously unrecognized biologic characteristics of platelets and are paralleled by new evidence for unique roles of platelets in inflammatory, immune, and thrombotic diseases.
Figures
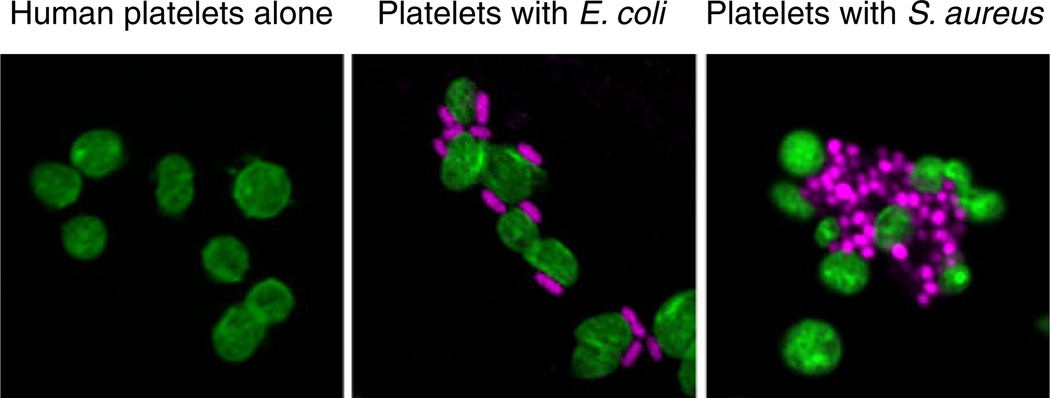
Human platelets recognize and interact with bacterial pathogens. Platelets (left panel) were incubated with E. coli (middle panel) or S. aureus (right panel) for 30 min in vitro followed by staining of platelet F-actin (green) and bacterial DNA (magenta) (from [149] with permission from the publisher). Platelets are sentinels that detect intravascular microbes of a variety of classes and respond with activities that contribute to the capture, containment, and elimination of pathogens and to complex innate and adaptive immune responses (Figs. 2 and 3). In some cases, platelet activation by bacteria or their toxins—for example, LPS released by E. coli or staphylococcal alpha toxin—leads to inflammatory vascular injury and thrombosis (see text and cited references for details)
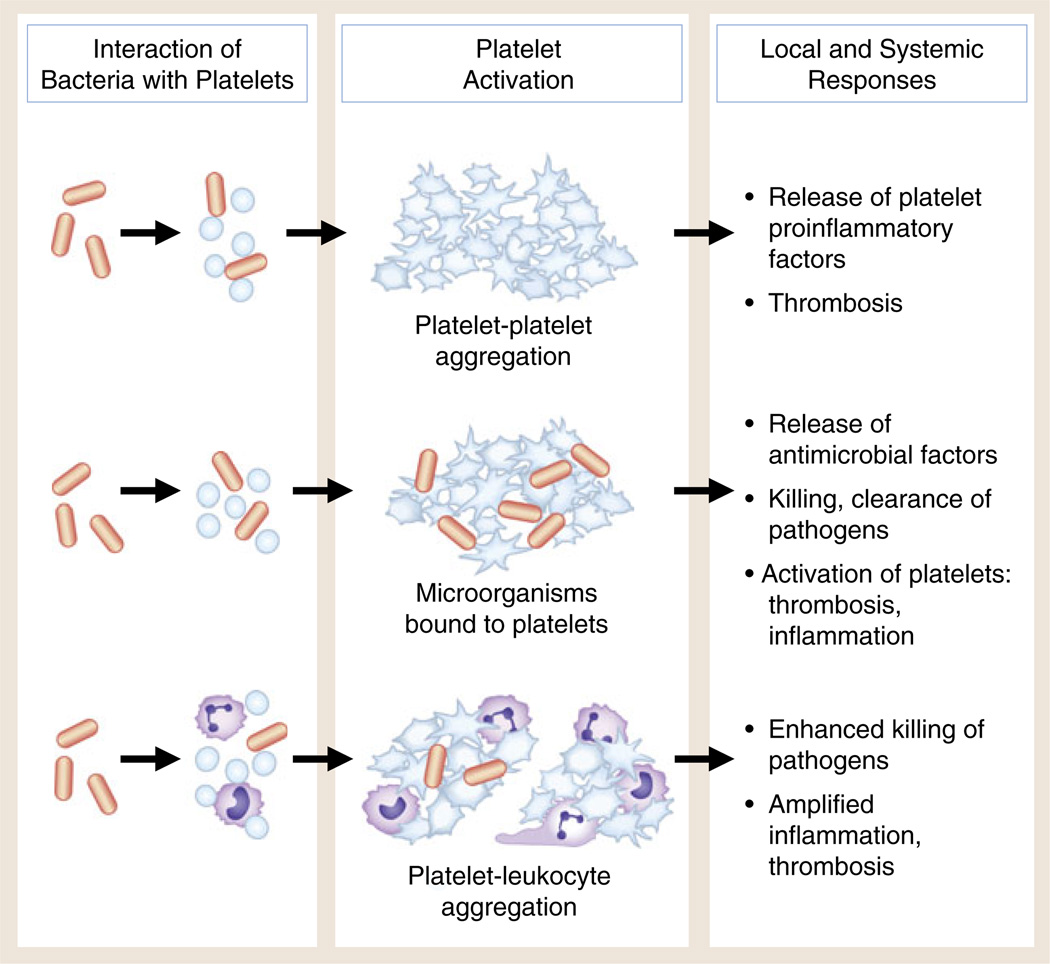
Interaction of bacteria with human platelets induces cellular activation and local and systemic thrombotic and inflammatory responses. Direct interaction of bacteria with platelets (see Fig. 1) can lead to aggregation, release of antimicrobial factors and proinflammatory molecules, and formation of platelet–leukocyte aggregates. Under some conditions, bacterial toxins of several classes can trigger these responses of human and murine platelets. Each functional activity is considered in detail in the text. There is evidence that these activities can mediate the containment, immobilization, and killing of bacteria and other pathogens. In addition, however, each can contribute to local thrombosis and inflammation in endovascular infectious syndromes such as bacterial endocarditis and to systemic infections with end-organ injury, including sepsis and complicated malaria
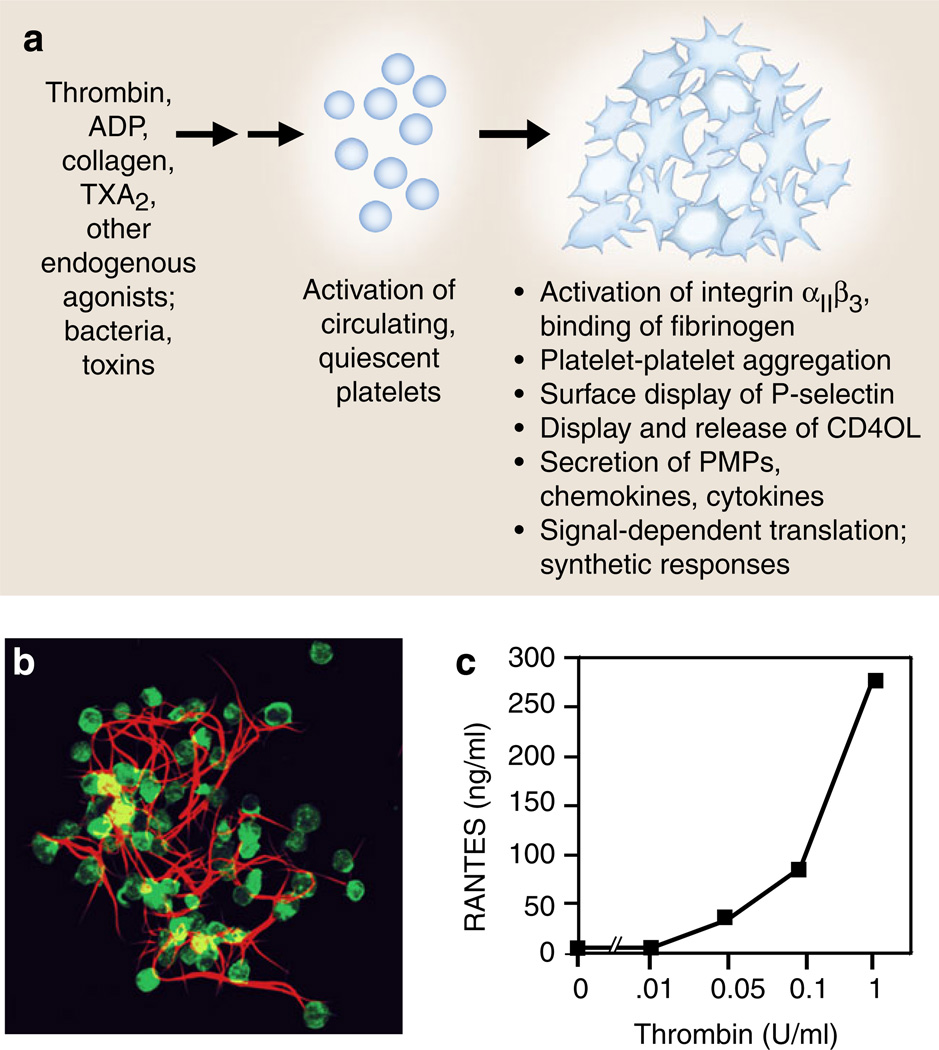
Activation responses of platelets mediate the critical events in hemostasis and inflammation. a Platelets activated by agonists recognized by G-protein-coupled receptors, including thrombin, ADP, and TxA2 or by immunoreceptors such as GPVI, a collagen receptor, deliver outside-in signals to human and murine platelets, resulting in rapid and, in some cases, sustained functional responses that mediate key events in hemostasis and inflammation, and often provide molecular links between hemostatic and immune pathways and responses. Activated platelets also release microvesicles that can initiate or amplify hemostasis and inflammation (see text for details). b Activated platelets bind fibrinogen via integrin αIIbβ3 and also bind other coagulation proteins, synthesize tissue factor, and promote the conversion of fibrinogen to fibrin as illustrated by this human platelet– fibrin clot formed in vitro. Activated platelets also retract fibrin, potentially consolidating and remodeling clots in vivo. The adhesive interactions of platelets with fibrinogen and fibrin can trigger additional activation and synthetic responses and inflammatory signaling (panel b is reproduced from [110] with permission from the publisher). c RANTES is an example of a preformed mediator with multiple inflammatory and immune activities that is secreted by activated human platelets (also see Table 1). In this experiment, RANTES was measured by ELISA in supernatants of human platelets activated with increasing concentrations of thrombin for 30 min as described in [171, 173] (panel c was modified from data in [173])
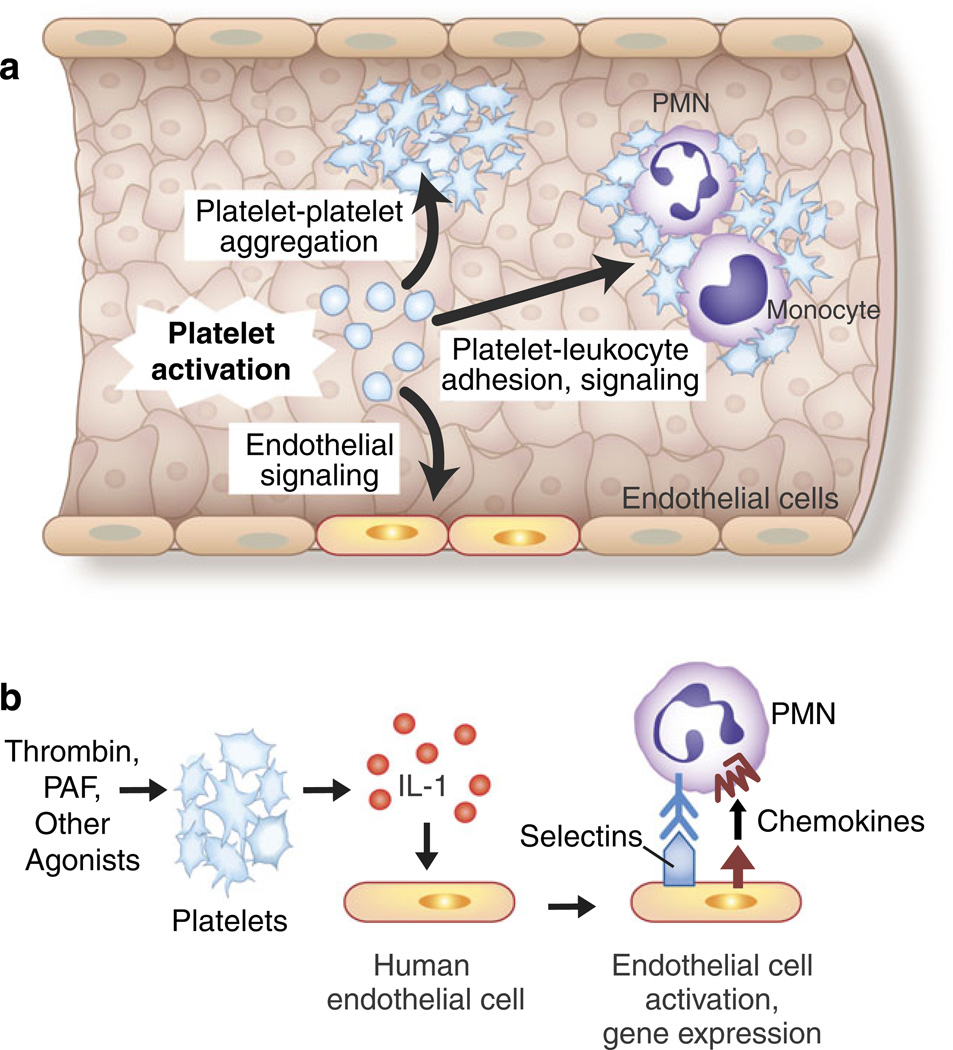
Adhesive and signaling mechanisms of activated platelets mediate critical cell–cell interactions in hemostasis, inflammation, and immune responses. a The activation of platelets mediates platelet– platelet aggregation, platelet–leukocyte adhesion and intercellular signaling, and interactions with inflamed and injured endothelial cells. Endothelial cells release factors that inhibit or modify these interactions, including prostacyclin and nitric oxide, under basal and stimulated conditions. The adhesion of platelets to exposed collagen and subendothelial matrix at sites of vascular injury is also a critical mechanism for platelet and leukocyte accumulation and for cell–cell interactions between platelets and leukocytes (not shown) (see text and Figs. 5, 6, 7, and 8 for additional details). b The release of IL-1 by activated platelets mediates platelet–endothelial interactions. The activated human and murine platelets release IL-1 in solution and associated with microvesicles. The release of both IL-1α and IL-1β has been reported, although in some studies IL-1 was only detected on the surfaces of activated platelets. Human endothelial cells release IL-8, MCP-1, IL-6, and GM-CSF and express E-selectin and increased levels of intercellular adhesion molecule 1 in response to juxtacrine signaling by IL-1 associated with activated platelets or IL-1 released by thrombin-stimulated platelets. IL-1β released in microvesicles from activated platelets also induces human endothelial cells to bind PMNs. This is consistent with many observations demonstrating that IL-1 is a prototypic agonist for “type II” activation of endothelial cells, which includes the synthesis and surface expression of E-selectin, IL-8, and other chemokines, leading to the adhesion and activation of PMNs and monocytes. IL-1 also induces the synthesis of tissue factor-dependent procoagulant activity by human endothelial cells. Thus, IL-1-mediated platelet-endothelial interactions have the potential to induce and drive key innate and adaptive immune responses and to link inflammation and hemostasis (see text, Table 1, and Fig. 5 for additional details and for references; Fig. 4 was adapted from [124])
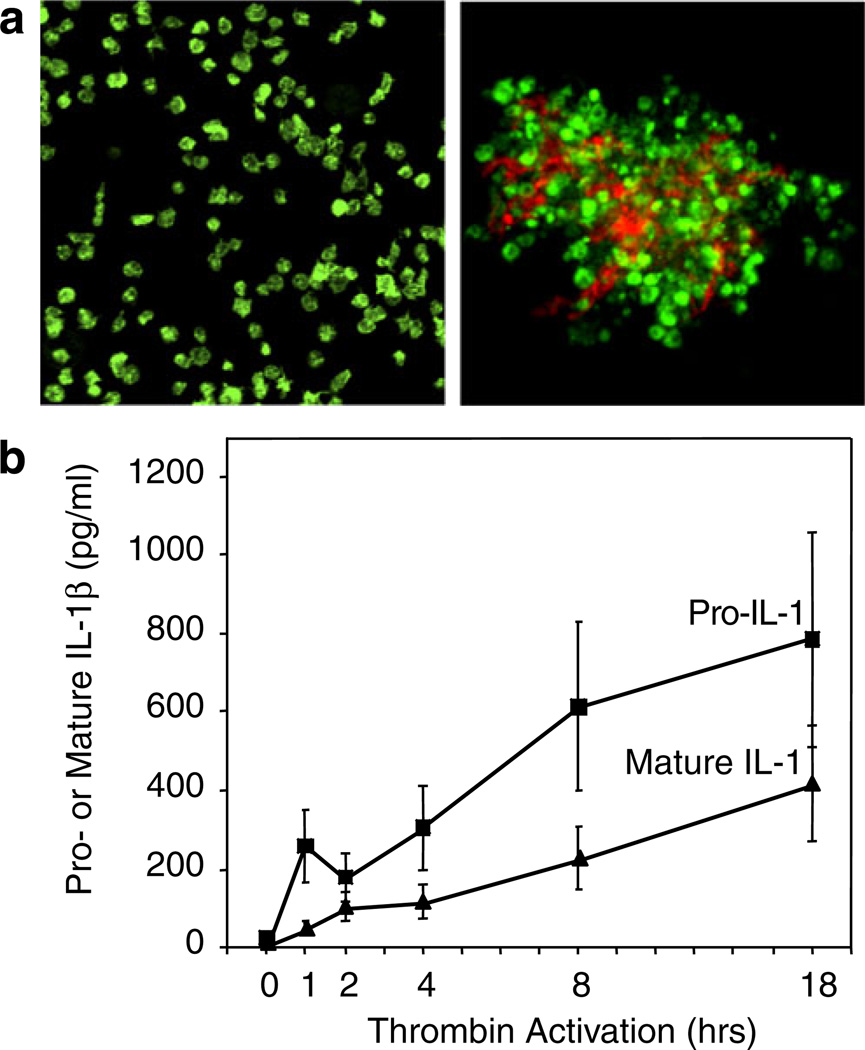
Activated human platelets synthesize IL-1β. IL-1β was not detected in freshly isolated unstimulated platelets by immunocyto-chemistry (aleft panel) or ELISA (b). Stimulation of platelets with thrombin (0.1 u/mL) in the presence of fibrinogen as a model of clot formation induced the rapid and sustained accumulation of the precursor and mature forms of IL-1β (b) that was blocked by the protein synthesis inhibitor puromycin [122]. Transcripts encoding IL-1β were present in resting, unactivated platelets and were found in polyribosomes of activated platelets. PAF, ADP, collagen, and epinephrine also induced IL-1β synthesis (not shown). IL-1β was detected in the fibrin lattice of platelet–fibrin complexes as well as associated with the platelets when platelet–fibrin complexes were examined by confocal microscopy (aright panel; IL-1β is stained in orange and platelet F-actin in green) and also in microvesicles shed from the activated platelets (not shown). Microvesicles from activated platelets induced cultured human endothelial monolayers to bind PMNs in an IL-1β-dependent fashion (Fig. 4b) (from [122] with permission from the publisher; see text and Fig. 9 for additional details of the mechanism of IL-1β synthesis)
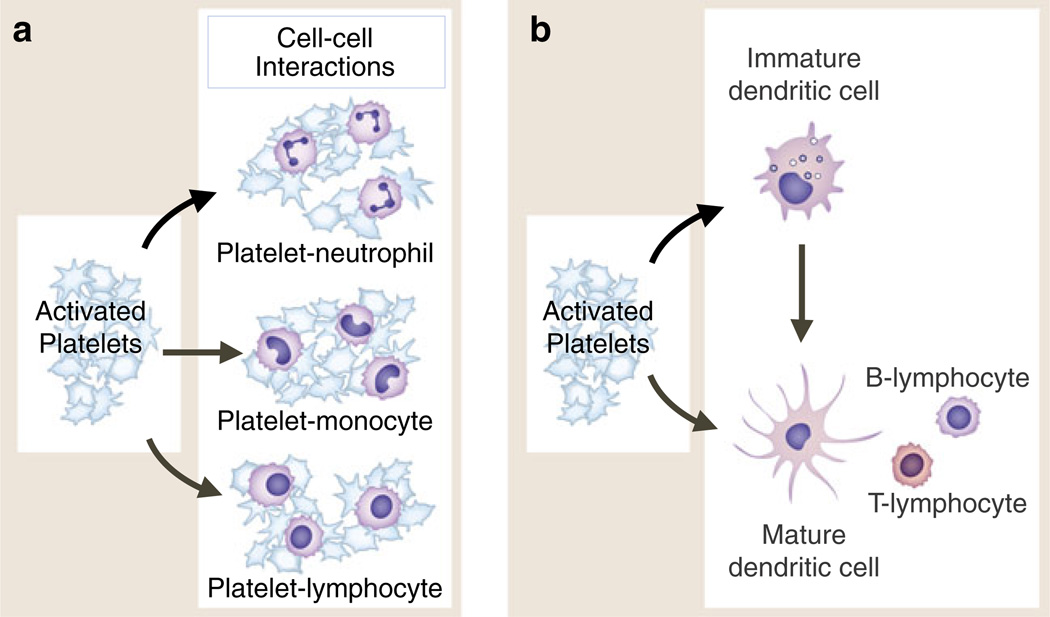
Activated platelets adhere to and signal circulating leukocytes and interact with dendritic cells and macrophages. Activated human platelets form heterotypic aggregates with neutrophils and other granulocytes (eosinophils, basophils), monocytes, and lymphocytes of several subclasses (a). In each interaction, binding of P-selectin on the activated platelet to PSGL-1 on the “target” leukocyte is critical. Additional adhesive mechanisms contribute to aggregate formation and to interactions with endothelial cells, subendothelial matrix, or other cell types depending on the specific leukocyte class involved. In addition to adhesion, intercellular signaling mediated by PSGL-1 engagement and other signal transduction pathways leads to altered functions of the leukocytes (see text and Fig. 7). Activated platelets also interact with and signal dendritic cells and, by these interactions, can indirectly alter lymphocyte activities (b). This may be a key mechanism by which activated platelets mediate information transfer between innate and adaptive components of the immune continuum and influence adaptive immune responses. Activated platelets interact with macrophages in addition to DC and may influence cell fate dermination when monocytes differentiate to immature dendritic cells or to macrophages (not shown). Activated platelets bind to monocytes and plasmacytoid DC in the circulation of patients with SLE and to PMNs and monocytes in a variety of other clinical conditions (see text for details)
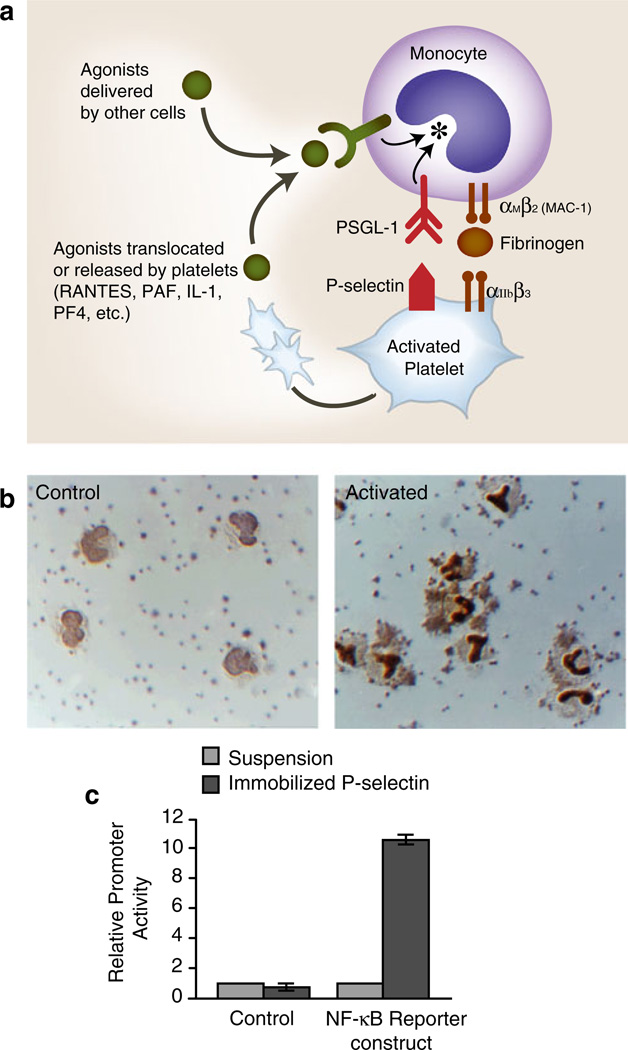
a Activated human platelets adhere to monocytes and deliver outside-in signals that induce altered functions. Activated human platelets adhere to monocytes in vivo and in vitro. Binding of P-selectin, which is translocated to the plasma membranes of activated platelets from alpha granules, to PSGL-1 basally present on the monocyte plasma membrane is critical for this interaction. The adhesion of activated platelets to monocytes is inhibited by blocking antibodies against P-selectin or PSGL-1 and by other approaches that interrupt this interaction. The engagement of PSGL-1 signals the activation of αMβ2and, likely, other members of the integrin family on monocytes, potentially amplifying the adhesion provided by P-selectin/PSGL-1 and contributing to other adhesive interactions with endothelium, subendothelial matrix, or other blood cells. Outside-in signals delivered by soluble or juxtacrine factors released or displayed by activated platelets (RANTES, PF4, IL-1, PAF, CD40L, etc.) can act in concert with signals delivered by the engagement of PSGL-1 (see Fig. 8) as can agonists delivered by other cell types. Signaling of monocytes by activated platelets alters transcriptional and post-transcriptional pathways in the leukocytes (see Fig. 8). b The activated human platelets induce the nuclear translocation of NF-kB in adherent monocytes. Under basal conditions in vitro, human platelets do not adhere to monocytes, and NF-kB is largely found in the monocyte cytoplasm (left panel, “control”). When platelets are activated with thrombin, platelet-monocyte aggregates rapidly form and NF-kB translocates to the nucleus as indicated by dark nuclear staining with an antibody against the p65 subunit of NF-kB. Both platelet-monocyte aggregate formation and NF-kB translocation are blocked by interrupting the binding of P-selectin on the activated platelets to PSGL-1 on the monocytes. In additional experiments, integrated signals delivered by platelet P-selectin and RANTES (see Fig. 3) were shown to induce the expression of the NF-kB-dependent gene products MCP-1 and IL-8 by the monocytes (see Fig. 8) (panel b is from [171] with permission from the publisher.) c Signaling by P-selectin induces NF-kB activation. U937 monocytic cells transfected with an NF-kB reporter or a control construct were incubated in suspension or allowed to adhere to purified, immobilized P-selectin and reporter activity was measured. Triggering of NF-kB reporter activity by adhesion to purified P-selectin is consistent with additional experiments, demonstrating that the engagement of PSGL-1 transmits outside-in signals to this pathway and that activated platelets deliver signals that alter patterns of inflammatory gene expression in monocytes (see also panels a, b, and Fig. 8; panel c is from [218] with permission from the publisher)
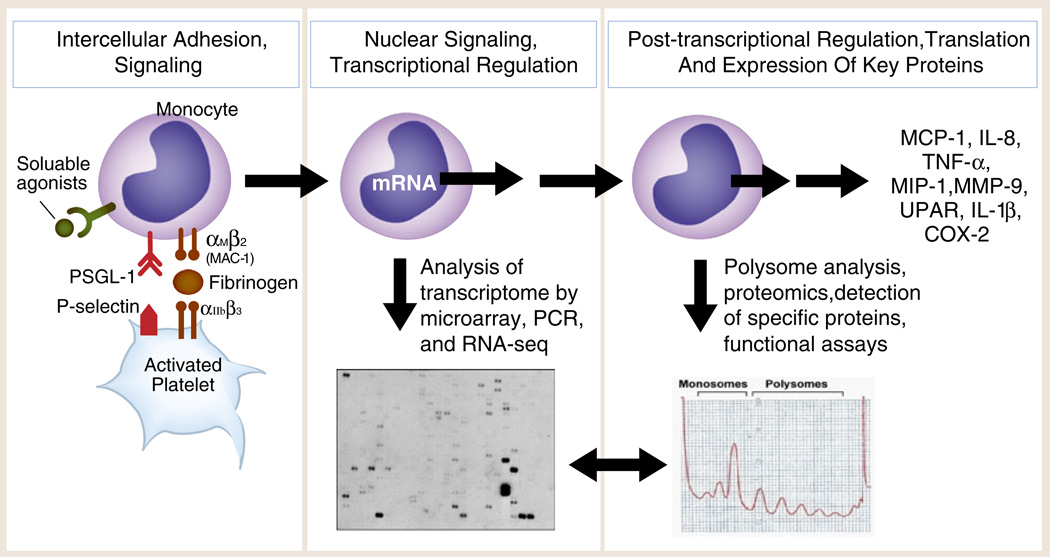
Outside-in signals delivered by activated, adherent human platelets induce the expression of inflammatory and thrombotic gene products by human monocytes. Experiments using thrombin-stimulated human platelets interacting with isolated monocytes as outlined in the left panel and in Fig. 7a demonstrate that activated platelets deliver outside-in signals to transcriptional (Fig. 7b, c) and post-transcriptional pathways (mRNA stabilization; mRNA association with RNA binding proteins; activation of mTOR, a regulator of specialized translation) via P-selectin/P-selectin engagement and parallel signaling by soluble or cell-associated platelet chemokines or cytokines. Assays of the monocyte transcriptome and transcrip-tional responses (middle panel) and proteomic analysis and functional assays (right panel) indicate that the protein products (far right) are synthesized by mechanisms that are time dependent and gene specific. Each protein product has important inflammatory and/or hemostatic activities based on many experimental and clinical studies. Two specific examples—expression of MCP-1 and COX-2—are outlined in the text (Fig. 8 was adapted from [277] with permission from the publisher)
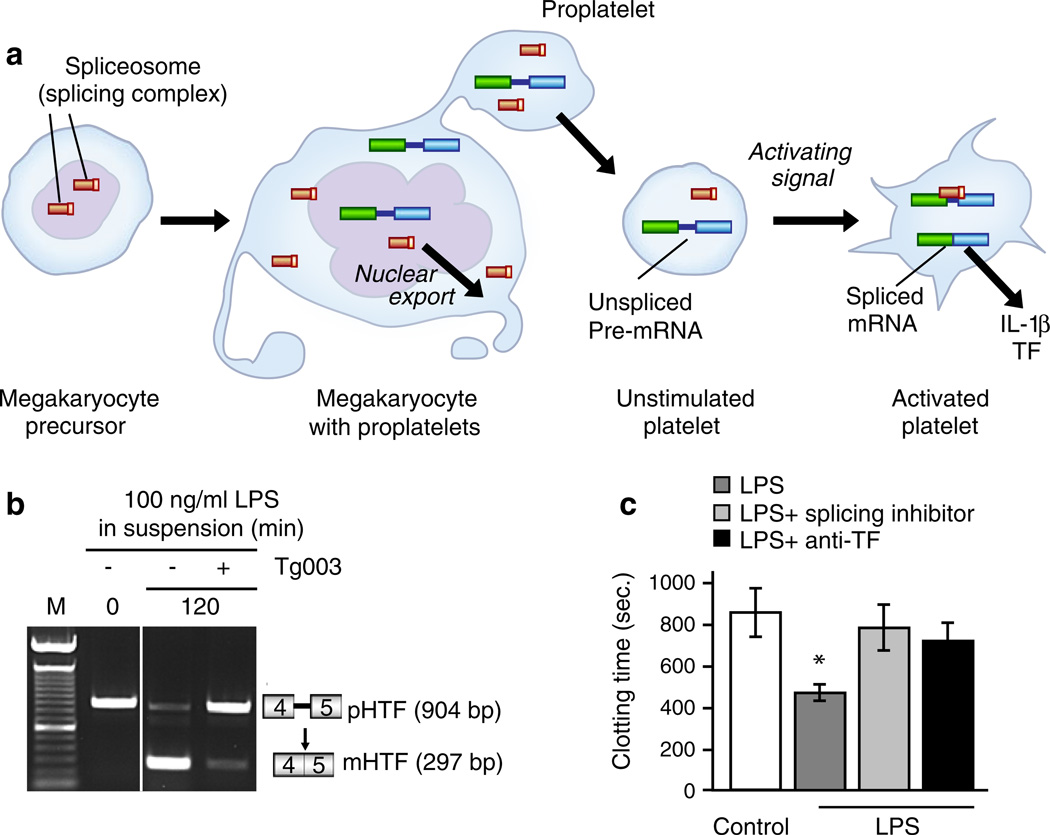
Activated human platelets synthesize inflammatory and hemostatic proteins by mechanisms involving signal-dependent translation and pre-mRNA splicing. An analysis of primary human platelets, cultured megakaryocytic cells, and proplatelet models demonstrate that human platelets have a complex transcriptome and post-transcriptional mechanisms for activation-dependent (“signal-dependent”) translation of mRNA transcripts [8]. One mechanism is illustrated in a: In addition to stable, mature mRNAs, the tran-scriptome of quiescent circulating human platelets includes unspliced pre-mRNAs. In response to thrombin or other activating signals, specific pre-mRNAs are processed by a functional spliceosome yielding the spliced, translatable mRNA. The pre-mRNAs and spliceosomal components are delivered to proplatelets during throm-bopoiesis, and an in situ analysis demonstrates the presence of unspliced pre-mRNAs in proplatelets and unstimulated platelets and in spliced transcripts in activated platelets. Following signal-dependent splicing, the mature, spliced mRNA is translated to biologically active protein. This pathway was first discovered in studies of the mechanism of synthesis of IL-1β by activated human platelets (Figs. 4b and 5) [14] (Fig. 9a is based on an illustration in [278]). Tissue factor is also synthesized by signal-dependent splicing and translation in platelets stimulated by thrombin and fibrinogen ([16]; not shown), bacteria (Fig. 1), or bacterial toxins. b A PCR analysis of the region of the TF pre-mRNA spanning exons 4 and 5 and the intervening intron (box diagrams at right of panel) revealed only the unspliced pre-mRNA in the control condition (time 0, lane 2; lane 1 indicates molecular weight markers). Treatment of the platelets with LPS for 120 min induced the conversion of the pre-mRNA to the spliced transcript that was nearly complete (lane 3). An inhibitor of Cdc2-like kinase 1, which is upstream in the splicing pathway, almost completely blocked the splicing and accumulation of the mature TF mRNA (lane 4). Parallel assays of TF-dependent procoagulant activity (c) demonstrated the shortening of the clotting time by LPS-stimulated platelets (bar 2 compared to bar 1; p<0.05) that was reversed by the splicing pathway inhibitor (bar 3) and an antibody against TF (bar 4). Additional analysis and controls can be found in [149] (panels b and c are from [149] with permission from the publisher). Additional pathways mediate signal-dependent translation of other mRNA transcripts by activated human platelets [8] (see text for details and additional references)
References
-
- Coller BS. A brief history of ideas about platelets in health and disease. In: Alan D, Michelson MD, editors. Platelets. 2nd edn. London: Elsevier; 2007. pp. xxiii–xlii.
-
- Weyrich AS, Lindemann S, Zimmerman GA. The evolving role of platelets in inflammation. J Thromb Haemost. 2003;1:1897–1905. - PubMed
-
- Weyrich AS, Zimmerman GA. Platelets: signaling cells in the immune continuum. Trends Immunol. 2004;25:489–495. - PubMed
Publication types
MeSH terms
Grants and funding
- ULI-RR025764/RR/NCRR NIH HHS/United States
- HL044525/HL/NHLBI NIH HHS/United States
- R01 HL066277/HL/NHLBI NIH HHS/United States
- HL066277/HL/NHLBI NIH HHS/United States
- R01 HL091754/HL/NHLBI NIH HHS/United States
- K231440921/PHS HHS/United States
- HL091754/HL/NHLBI NIH HHS/United States
- R01 HL044525/HL/NHLBI NIH HHS/United States
- HL092746/HL/NHLBI NIH HHS/United States
- R01 HL048872/HL/NHLBI NIH HHS/United States
- R01 HL090870/HL/NHLBI NIH HHS/United States
- HL048872/HL/NHLBI NIH HHS/United States
- HL090870/HL/NHLBI NIH HHS/United States
- R37 HL044525/HL/NHLBI NIH HHS/United States
- C06 RR011234/RR/NCRR NIH HHS/United States
- R01 HL092746/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
