

Review
doi: 10.1002/dneu.20962.
Building the posterior lateral line system in zebrafish
Affiliations
- PMID: 21818862
- PMCID: PMC3376715
- DOI: 10.1002/dneu.20962
Item in Clipboard
Review
Building the posterior lateral line system in zebrafish
Dev Neurobiol.
2012 Mar.
Abstract
The posterior lateral line (pLL) in zebrafish has emerged as an excellent system to study how a sensory organ system develops. Here we review recent studies that illustrate how interactions between multiple signaling pathways coordinate cell fate,morphogenesis, and collective migration of cells in the posterior lateral line primordium. These studies also illustrate how the pLL system is contributing much more broadly to our understanding of mechanisms operating during the growth, regeneration, and self-organization of other organ systems during development and disease.
Figures

Periodic deposition of neuromasts by the posterior Lateral Line Primordium (pLLP) A. Schematic of migration of the pLLP from the otic vesicle to the tip of the tail as it periodically deposits primary neuromasts L1 to L8. B. Images from a time lapse movie made using confocal microscopy of pLLP migration using a CldnB::LynGFP transgenic embryo showing progressive stages of neuromast deposition. Arrow points to depositing neuromast. C. Superimposition of a bright field and fluorescent image of a zebrafish larva at 5 days with neuromasts labelled with Yo-Pro-1. Arrows point to primary neuromasts L1 to L8 on the left side of the embryo, the image shows secondary neuromasts (*) and a few neuromasts from the right side of the embryo (not labelled).

Polarized Wnt and FGF signaling in the pLLP. A. The leading end of the pLLP is dominated by Wnt signaling while FGF signaling dominates at the trailing end. Cells assume a mesenchymal morphology at the leading end, where Wnt signaling dominates, while FGF signaling facilitates adoption of epithelial morphology and formation of epithelial rosettes, in trailing domains. B. Two color whole mount in situ hybridization showing Wnt signaling-dependent lef1 expression in the leading domain (red brown) and FGF signaling–dependent pea3 expression (purple) in the trailing domain, with some area of overlap in between. C. A Wnt-dependent source of FGF3 and FGF10 at the leading end establishes a domain of FGF signaling in an adjacent domain. While Wnt signaling drives expression of Sef, which locally inhibits FGF signaling, FGF signaling drives expression of Dkk1, which inhibits Wnt signaling.

Wnt-dependent FGF signaling facilitates morphogenesis of an epithelial rosette in a newly formed proto-neuromast. A. A Wnt-dependent source of FGFs (red cells) establishes a domain of FGF signaling (green cells) in an adjacent domain. B. FGF signaling in trailing cells facilitates adoption of epithelial morphology and apical constriction, which contributes to epithelial rosette formation.

Wnt-dependent FGF signaling initiates center biased atoh1a expression in a newly formed proto-neuromast (A). B. FGF-dependent pea3 expression reveals restricted center-biased FGF signaling at the leading end of the pLLP (arrow). This initiates center biased atoh1a and deltaA expression. Atoh1a initiates deltaD expression and together DeltaA and DeltaD activate Notch in neighboring cells, which inhibits atoh1a expression. C. Lateral inhibition, mediated by Notch signaling keeps atoh1a restricted to a central cell. This cell becomes a hair cell progenitor, while its neighbors, in which atoh1a is supressed become support cells. When Notch signaling fails atoh1a expression spreads to adjacent cells and hair cell progenitors (blue) expand in number at the cost of support cells (orange).
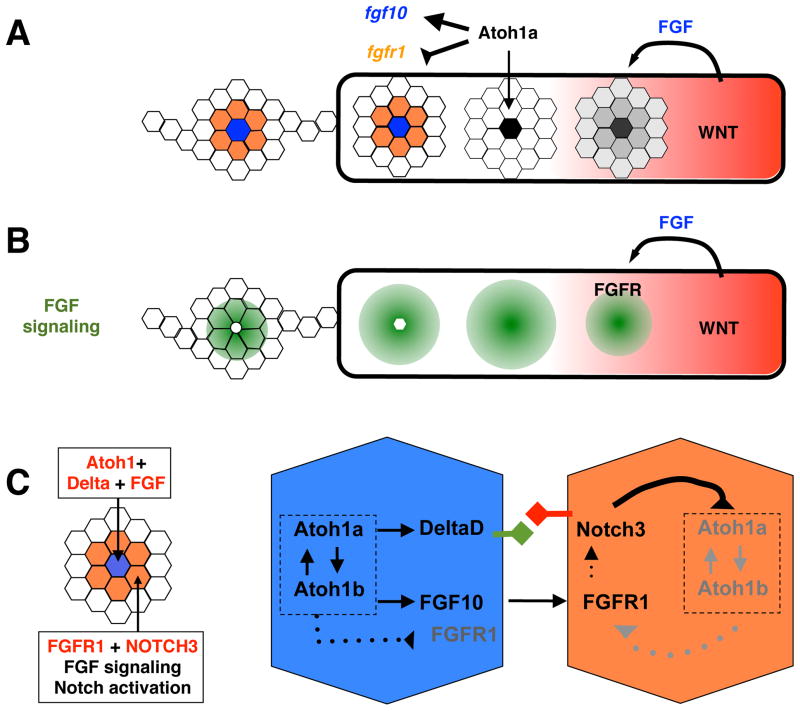
The central atoh1a-expressing cell becomes a new source of FGF10 in maturing proto-neuromasts. A. Atoh1a drives expression of fgf10 and inhibits expression of fgfr1. B. As a source of FGF10, the atoh1a expressing cell maintains FGF signaling at the trailing end of the pLLP. However, loss of fgfr1 expression makes this cell less responsive to FGF signaling. C. In the most mature trailing proto-neuromast Atoh1a drives expression of atoh1b, Atoh1b drives atoh1a expression. In this manner cross-activation helps maintain atoh1a expresion in trailing neuromasts, where its expression becomes less dependent on FGF signaling. As the central cell becomes a source of DeltaD and FGF10 it maintains Notch and FGF signaling in neighboring cells. As long as Notch activation is maintained in the neighboring cells they do not express atoh1a in response to FGF signaling.

A speculative model of how support cells expressing both e-cadherin and n-cadherin contribute to cohesive integrity of the pLL system A. (a) Comparative expression of atoh1a (black), e-cadherin (green), n-cadherin (red) in neuromast cells with distinct fate in a wild-type (WT) and mib1 mutant (No Notch) embryo. (b) A side view illustrates how expression of both e-cadherin (green) and n-cadherin in support cells (yellow) would allow them to develop adhesive junctions (arrows) with both central n-cadherin expressing hair cell progenitors (red) and surrounding non-sensory cells expressing only e-cadherin (green). (c) In the absence of Notch signaling hair cell progenitors expand at the expense of support cells, (d) resulting in a loss of cohesive interactions. B. (a) Schematic of the relatively complementary expression n-cadherin (red) and e-cadherin (green) in the migrating the pLLP. Prospective support cells in the trailing domain (yellow) express both e- and n-cadherin. (b) Loss of prospective support cells could contribute to fragmentation of the pLLP.

Polarized expression of CXCR4b and CXCR7b in the pLLP. A. Degradation of CXCL12a by CXCR7b may contribute to establishment of a transient local gradient of CXCL12a. Failure of CXCR7b activation in trailing cells to promote protrusive migratory activity ensures a polarized response to CXCL12a expressed in a relatively pattern in the migratory path. B. Schematic of potential interactions that determine polarized chemokine receptor expression. C. Trailing epithelial cells contribute to collective migration by extending crptic basal lamellipodia.

An evolving model of the neuromast deposition cycle. (a–d) Progressive changes in the pLLP during a deposition cycle. The leading cohesive column of migrating cxcr7-free cells is shown in green. Trailing cxcr7-expressing cells slowing down for deposition are shown in red. The size bar shows length of the migrating column of green cells remains relatively constant as proliferation adds to the total length of the migrating pLLP. In b, the dotted lines show formation of a new proneuomast as the pLLP lengthens. Prospective and deposited neuromasts are serially labelled 1–4 in the schematic.

The anatomy and organization of sensory hair cells. A. The central hair cell progenitor (blue) divides to form two hair cells. Microtuble-containing kinocilia form at eccentric adjacent positions in daughter cells, along an axis defined by the direction of migration of the pLLP prior to neuromast depsoition. Development of the kinocilium is accompanied by formation of rows of progressively shorter actin containing stereocilia. B. Schematic of a side view of a hair cell. The tip of each process is attached to the adjacent taller process by a linking filament. Tension in these links influences the probability of cation channels opening. Hence bending toward the kinocilum increases probability of channel opening while bending away contributes to lower probability of opening. C. During regeneration a hair cell progenitor divides to form daughter with opposing polarity. (i) A new hair cell progenitor is then specified toward either dorsal or ventral poles of the neuromast. (ii) In this manner adjacent columns of hair cells with opposing planar plolarity are progressively established. Eventually the ellipsoid shape of the neuromast is lost and it become more rounded as hair cells move away from the equator to establish mirror symmetric populations with opposing planar polarity.
References
-
- Aman A, Nguyen M, Piotrowski T. Wnt/beta-catenin dependent cell proliferation underlies segmented lateral line morphogenesis. Dev Biol. 2011;349:470–482. - PubMed
-
- Aman A, Piotrowski T. Wnt/beta-catenin and Fgf signaling control collective cell migration by restricting chemokine receptor expression. Dev Cell. 2008;15:749–761. - PubMed
-
- Baker PC, Schroeder TE. Cytoplasmic filaments and morphogenetic movement in the amphibian neural tube. Developmental Biology. 1967;15:432–450. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
