

Optogenetic control of cells and circuits
- PMID: 21819234
- PMCID: PMC3759011
- DOI: 10.1146/annurev-cellbio-100109-104051
Optogenetic control of cells and circuits
Abstract
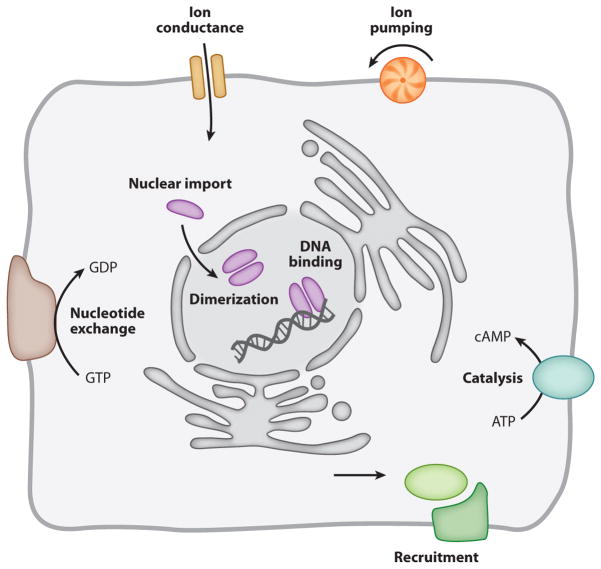
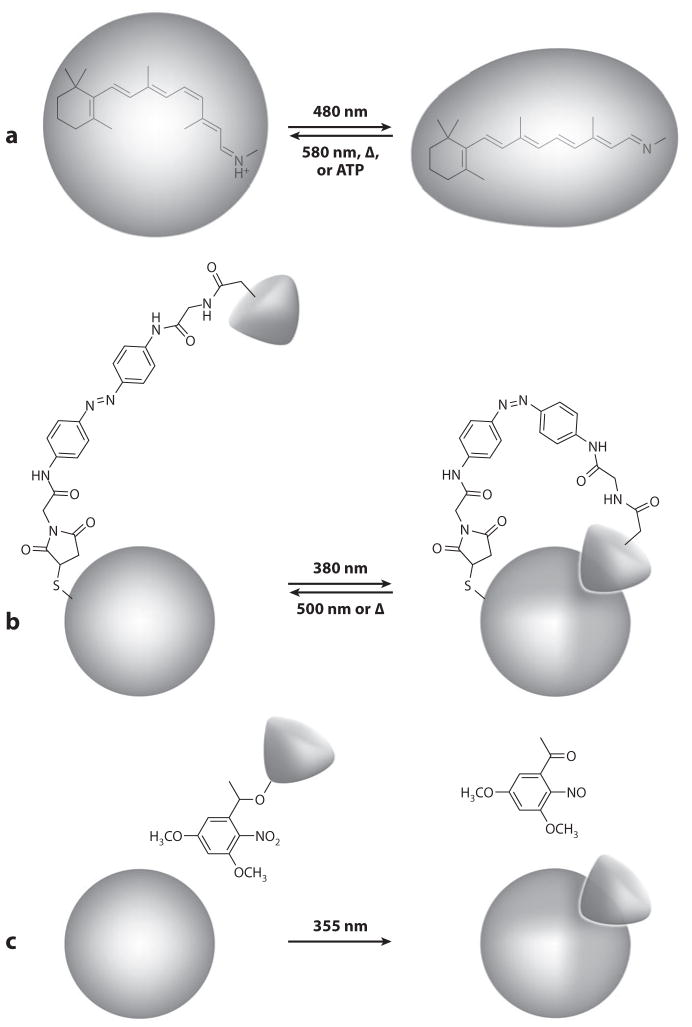
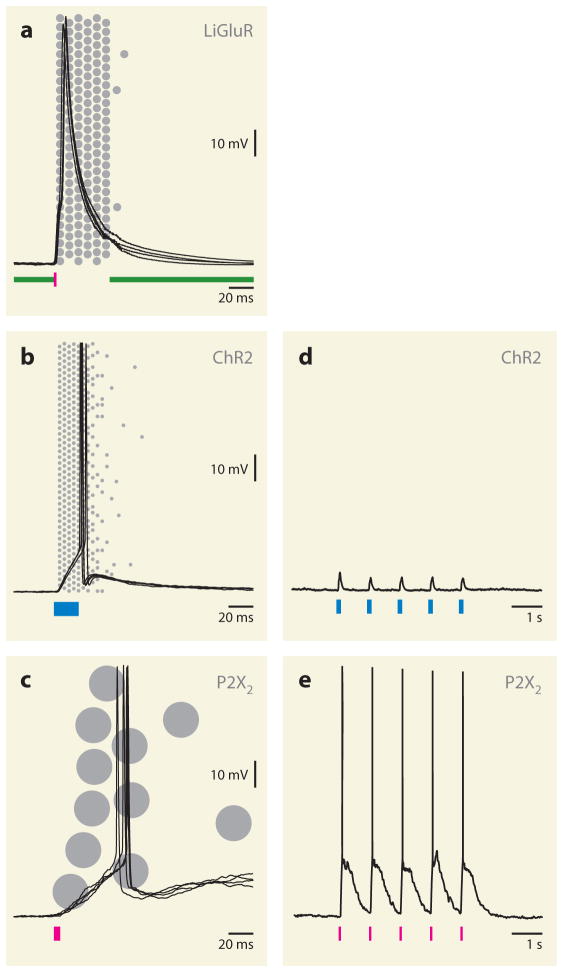
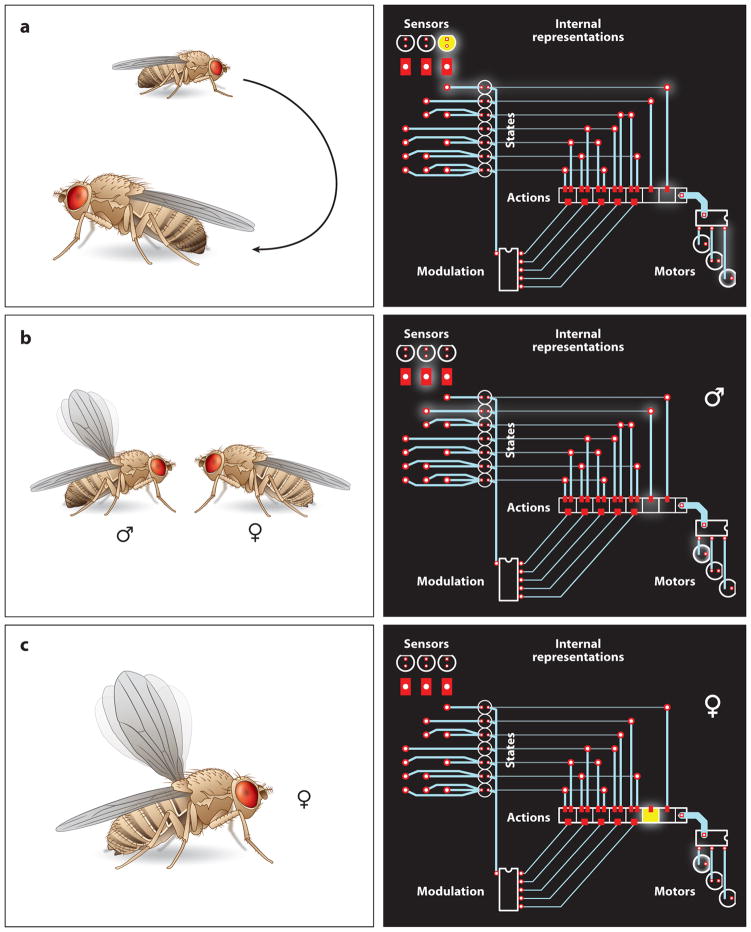
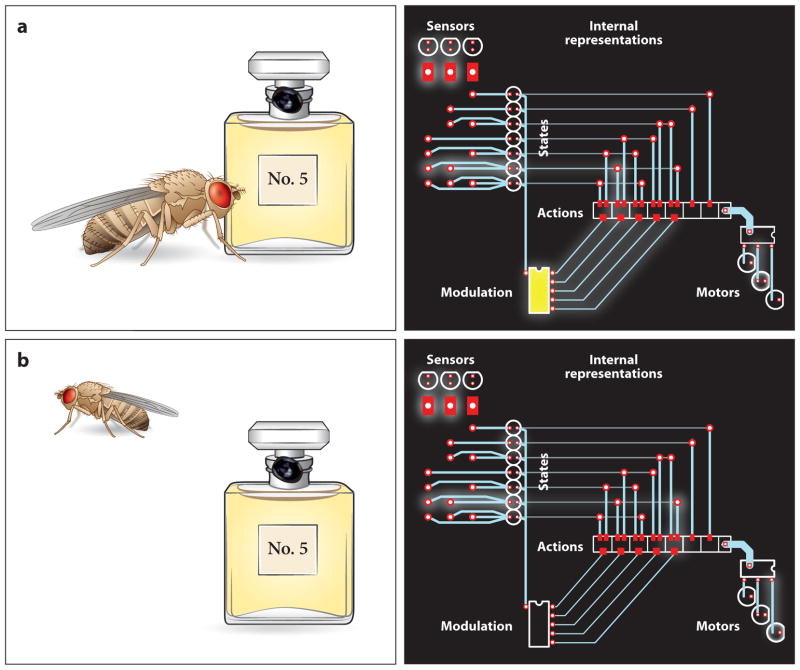
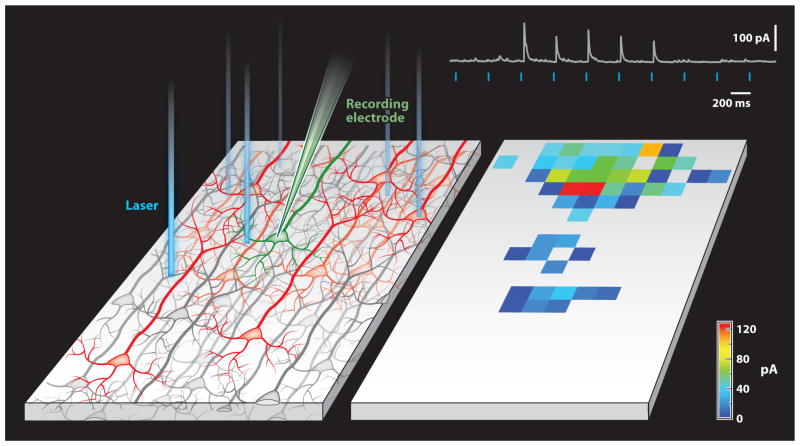
The absorption of light by bound or diffusible chromophores causes conformational rearrangements in natural and artificial photoreceptor proteins. These rearrangements are coupled to the opening or closing of ion transport pathways, the association or dissociation of binding partners, the enhancement or suppression of catalytic activity, or the transcription or repression of genetic information. Illumination of cells, tissues, or organisms engineered genetically to express photoreceptor proteins can thus be used to perturb biochemical and electrical signaling with exquisite cellular and molecular specificity. First demonstrated in 2002, this principle of optogenetic control has had a profound impact on neuroscience, where it provides a direct and stringent means of probing the organization of neural circuits and of identifying the neural substrates of behavior. The impact of optogenetic control is also beginning to be felt in other areas of cell and organismal biology.
Figures
References
-
- Adams SR, Tsien RY. Controlling cell chemistry with caged compounds. Annu Rev Physiol. 1993;55:755–84. - PubMed
-
- Airan RD, Thompson KR, Fenno LE, Bernstein H, Deisseroth K. Temporally precise in vivo control of intracellular signalling. Nature. 2009;458:1025–29. - PubMed
-
- Aravanis AM, Wang LP, Zhang F, Meltzer LA, Mogri MZ, et al. An optical neural interface: in vivo control of rodent motor cortex with integrated fiberoptic and optogenetic technology. J Neural Eng. 2007;4:S143–56. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
