

Exploring the roles of nucleobase desolvation and shape complementarity during the misreplication of O(6)-methylguanine
- PMID: 21819995
- PMCID: PMC3167027
- DOI: 10.1016/j.jmb.2011.07.011
Exploring the roles of nucleobase desolvation and shape complementarity during the misreplication of O(6)-methylguanine
Abstract
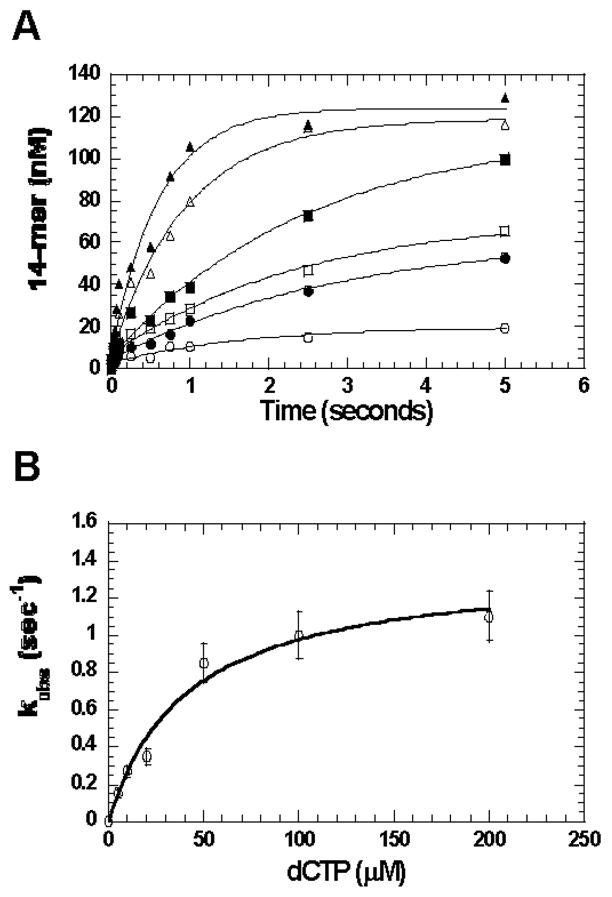
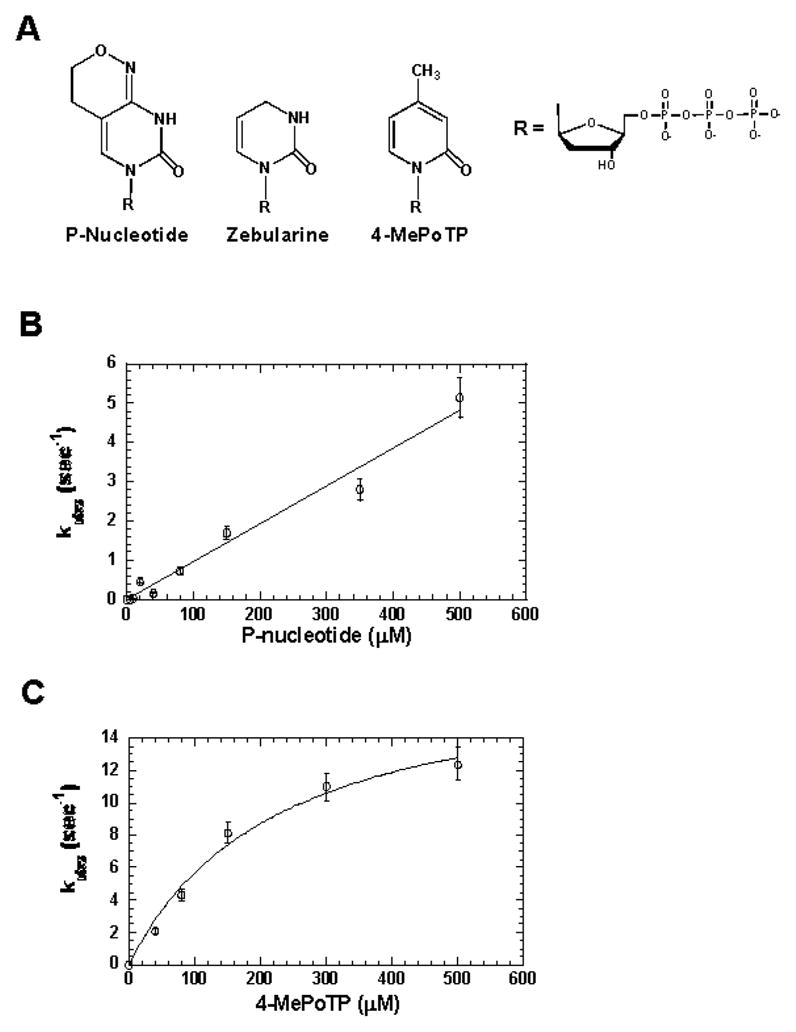
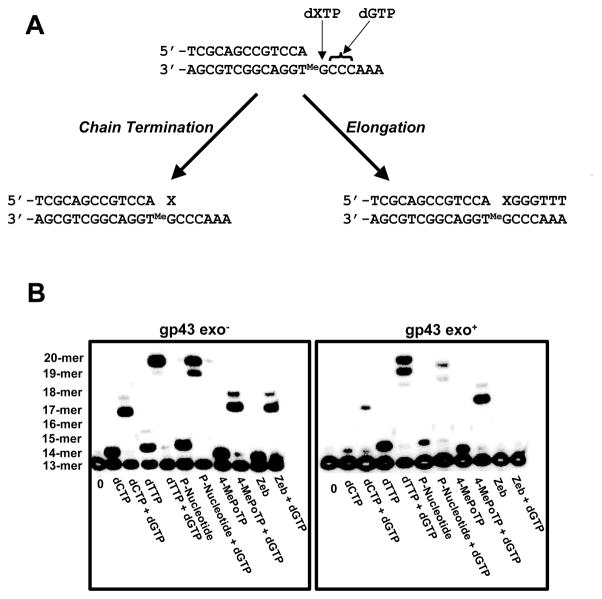
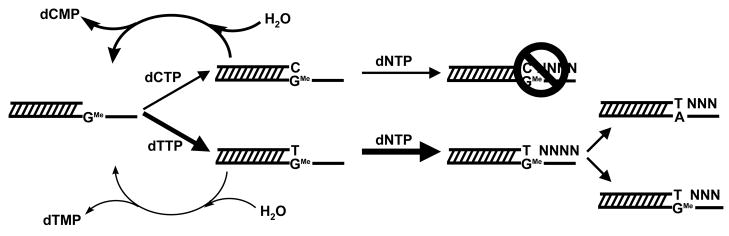
O(6)-methylguanine (O(6)-MeG) is a miscoding DNA lesion arising from the alkylation of guanine. This report uses the bacteriophage T4 DNA polymerase as a model to probe the roles of hydrogen-bonding interactions, shape/size, and nucleobase desolvation during the replication of this miscoding lesion. This was accomplished by using transient kinetic techniques to monitor the kinetic parameters for incorporating and extending natural and nonnatural nucleotides. In general, the efficiency of nucleotide incorporation does not depend on the hydrogen-bonding potential of the incoming nucleotide. Instead, nucleobase hydrophobicity and shape complementarity appear to be the preeminent factors controlling nucleotide incorporation. In addition, shape complementarity plays a large role in controlling the extension of various mispairs containing O(6)-MeG. This is evident as the rate constants for extension correlate with proper interglycosyl distances and symmetry between the base angles of the formed mispair. Base pairs not conforming to an acceptable geometry within the polymerase's active site are refractory to elongation and are processed via exonuclease proofreading. The collective data set encompassing nucleotide incorporation, extension, and excision is used to generate a model accounting for the mutagenic potential of O(6)-MeG observed in vivo. In addition, kinetic studies monitoring the incorporation and extension of nonnatural nucleotides identified an analog that displays high selectivity for incorporation opposite O(6)-MeG compared to unmodified purines. The unusual selectivity of this analog for replicating damaged DNA provides a novel biochemical tool to study translesion DNA synthesis.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures




Similar articles
-
Evaluating the contribution of base stacking during translesion DNA replication.Biochemistry. 2004 Jan 20;43(2):393-404. doi: 10.1021/bi034948s. Biochemistry. 2004. PMID: 14717593
-
Spectroscopic analysis of polymerization and exonuclease proofreading by a high-fidelity DNA polymerase during translesion DNA synthesis.Biochim Biophys Acta. 2013 Jan;1834(1):34-45. doi: 10.1016/j.bbapap.2012.08.019. Epub 2012 Aug 28. Biochim Biophys Acta. 2013. PMID: 22959853
-
Molecular basis of selectivity of nucleoside triphosphate incorporation opposite O6-benzylguanine by sulfolobus solfataricus DNA polymerase Dpo4: steady-state and pre-steady-state kinetics and x-ray crystallography of correct and incorrect pairing.J Biol Chem. 2007 May 4;282(18):13573-84. doi: 10.1074/jbc.M700656200. Epub 2007 Mar 3. J Biol Chem. 2007. PMID: 17337730
-
The steric hypothesis for DNA replication and fluorine hydrogen bonding revisited in light of structural data.Acc Chem Res. 2012 Aug 21;45(8):1237-46. doi: 10.1021/ar200303k. Epub 2012 Apr 23. Acc Chem Res. 2012. PMID: 22524491 Free PMC article. Review.
-
RB69 DNA polymerase structure, kinetics, and fidelity.Biochemistry. 2014 May 6;53(17):2752-67. doi: 10.1021/bi4014215. Epub 2014 Apr 23. Biochemistry. 2014. PMID: 24720884 Free PMC article. Review.
Cited by
-
The use of an artificial nucleotide for polymerase-based recognition of carcinogenic O6-alkylguanine DNA adducts.Nucleic Acids Res. 2016 Aug 19;44(14):6564-73. doi: 10.1093/nar/gkw589. Epub 2016 Jul 4. Nucleic Acids Res. 2016. PMID: 27378785 Free PMC article.
-
Development of a 'clickable' non-natural nucleotide to visualize the replication of non-instructional DNA lesions.Nucleic Acids Res. 2012 Mar;40(5):2357-67. doi: 10.1093/nar/gkr980. Epub 2011 Nov 15. Nucleic Acids Res. 2012. PMID: 22086959 Free PMC article.
-
Incorporation of nucleoside probes opposite O⁶-methylguanine by Sulfolobus solfataricus DNA polymerase Dpo4: importance of hydrogen bonding.Chembiochem. 2013 Sep 2;14(13):1634-9. doi: 10.1002/cbic.201300296. Epub 2013 Aug 19. Chembiochem. 2013. PMID: 23959784 Free PMC article.
-
DNA damage by reactive species: Mechanisms, mutation and repair.J Biosci. 2012 Jul;37(3):503-17. doi: 10.1007/s12038-012-9218-2. J Biosci. 2012. PMID: 22750987 Review.
References
-
- Kunkel TA, Bebenek K. DNA replication fidelity. Annu Rev Biochem. 2000;69:497–529. - PubMed
-
- Grevatt PC, Solomon JJ, Bhanot OS. In vitro mispairing specificity of O2-ethylthymidine. Biochemistry. 1992;31:4181–4188. - PubMed
-
- Marcelino LA, Andre PC, Khrapko K, Coller HA, Griffith J, Thilly WG. Chemically induced mutations in mitochondrial DNA of human cells: mutational spectrum of N-methyl-N′-nitro-N-nitrosoguanidine. Cancer Res. 1998;58:2857–2862. - PubMed
-
- Yan SF, Wu M, Geacintov NE, Broyde S. Altering DNA polymerase incorporation fidelity by distorting the dNTP binding pocket with a bulky carcinogen-damaged template. Biochemistry. 2004;43:7750–7765. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
