

Mechanisms of atrial-selective block of Na⁺ channels by ranolazine: I. Experimental analysis of the use-dependent block
- PMID: 21821778
- PMCID: PMC3197375
- DOI: 10.1152/ajpheart.00242.2011
Mechanisms of atrial-selective block of Na⁺ channels by ranolazine: I. Experimental analysis of the use-dependent block
Abstract
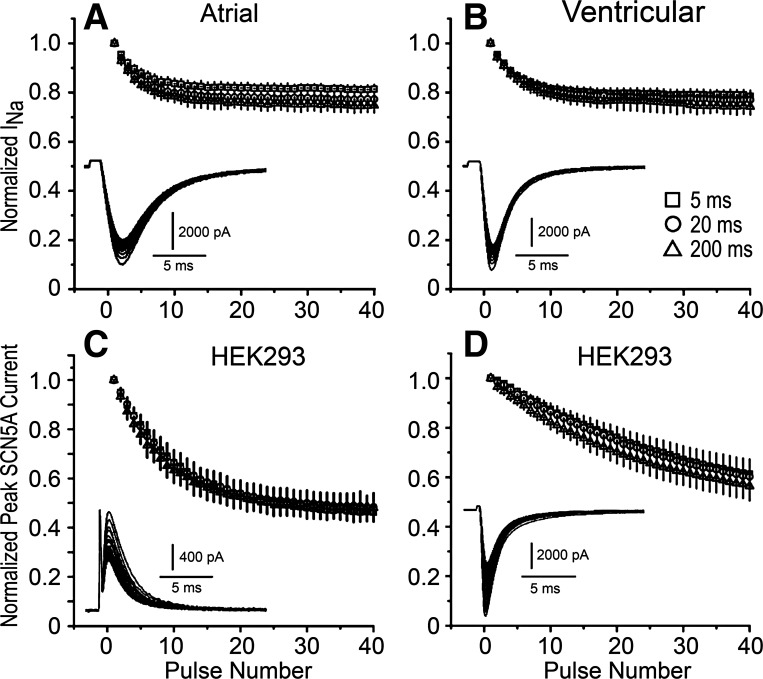
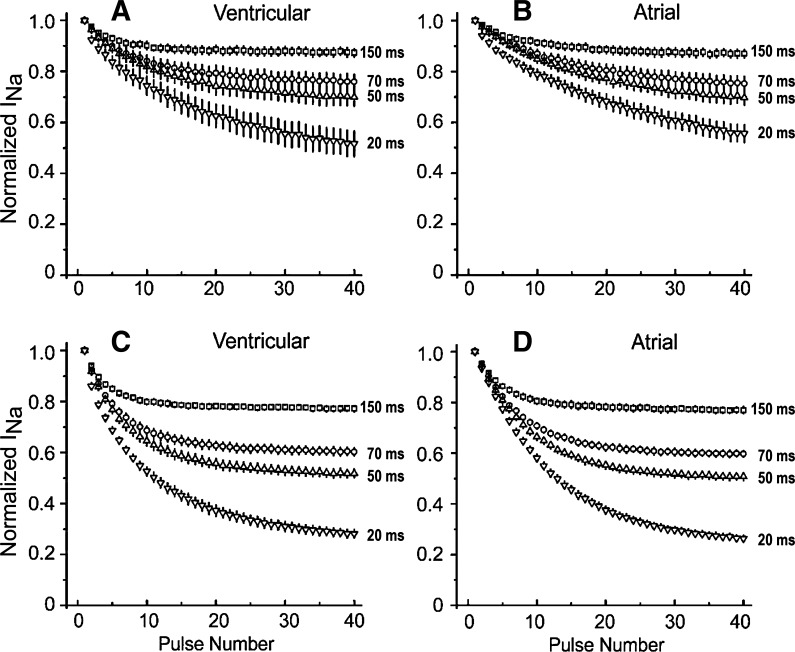
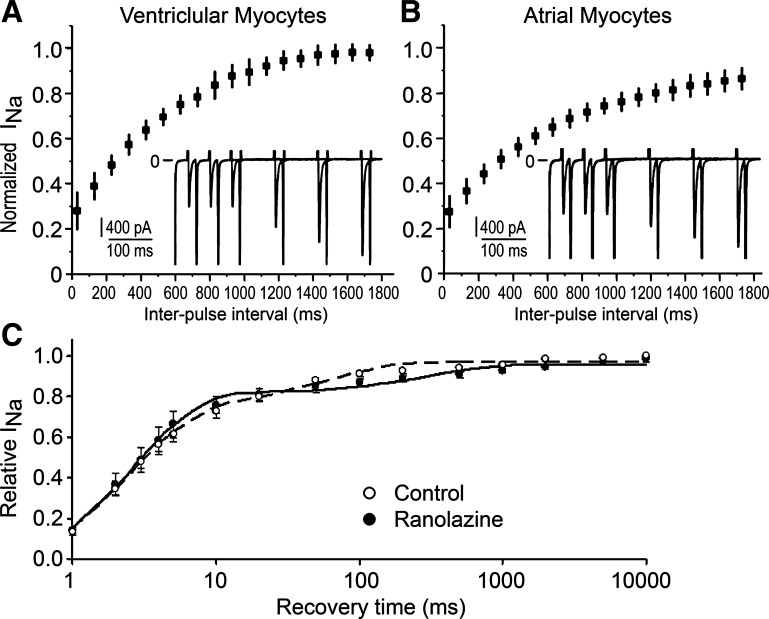
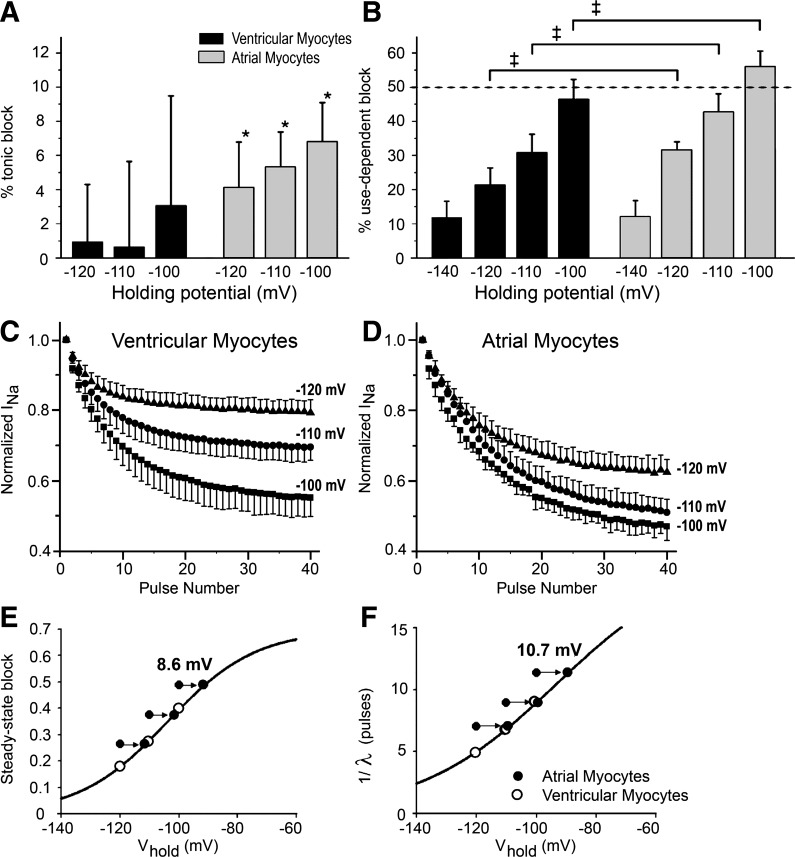
Atrial-selective inhibition of cardiac Na(+) channel current (I(Na)) and I(Na)-dependent parameters has been shown to contribute to the safe and effective management of atrial fibrillation. The present study examined the basis for the atrial-selective actions of ranolazine. Whole cell I(Na) was recorded at 15°C in canine atrial and ventricular myocytes and in human embryonic kidney (HEK)-293 cells expressing SCN5A. Tonic block was negligible at holding potentials from -140 to -100 mV, suggesting minimal drug interactions with the closed state. Trains of 40 pulses were elicited over a range of holding potentials to determine use-dependent block. Guarded receptor formalism was used to analyze the development of block during pulse trains. Use-dependent block by ranolazine increased at more depolarized holding potentials, consistent with an interaction of the drug with either preopen or inactivated states, but was unaffected by longer pulse durations between 5 and 200 ms, suggesting a weak interaction with the inactivated state. Block was significantly increased at shorter diastolic intervals between 20 and 200 ms. Responses in atrial and ventricular myocytes and in HEK-293 cells displayed a similar pattern. Ranolazine is an open state blocker that unbinds from closed Na(+) channels unusually fast but is trapped in the inactivated state. Kinetic rates of ranolazine interactions with different states of atrial and ventricular Na(+) channels were similar. Our data suggest that the atrial selectivity of ranolazine is due to a more negative steady-state inactivation curve, less negative resting membrane potential, and shorter diastolic intervals in atrial cells compared with ventricular cells at rapid rates.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous