

Nucleophosmin deposition during mRNA 3' end processing influences poly(A) tail length
- PMID: 21822216
- PMCID: PMC3209774
- DOI: 10.1038/emboj.2011.272
Nucleophosmin deposition during mRNA 3' end processing influences poly(A) tail length
Abstract
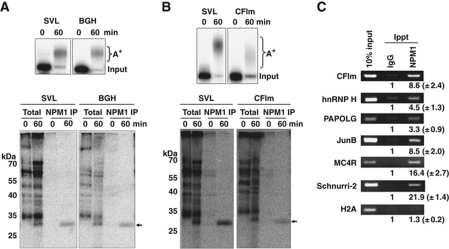
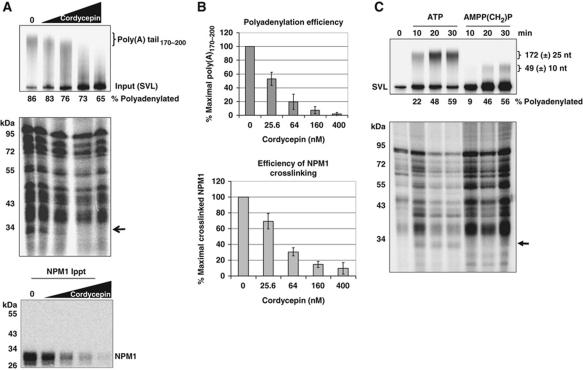
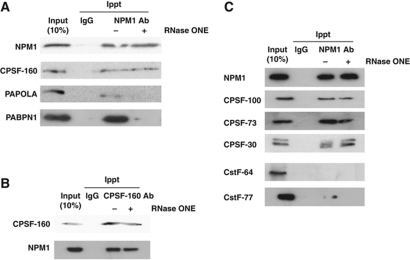
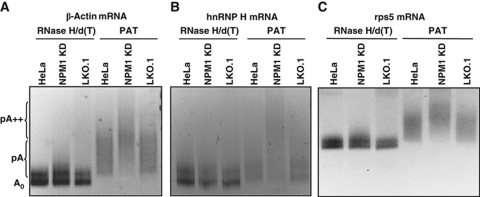
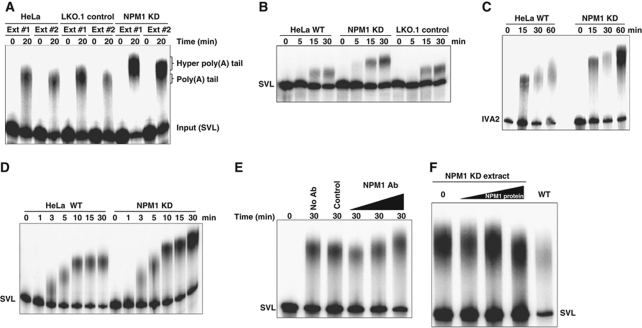
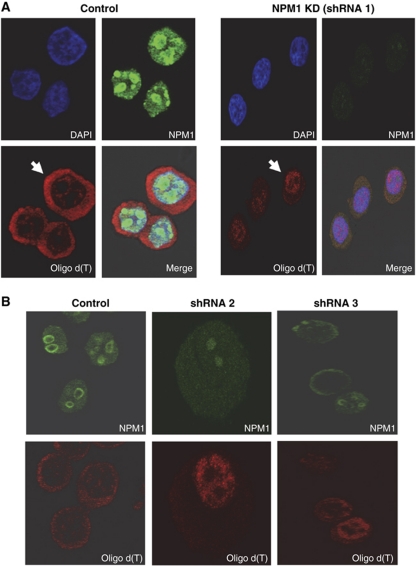
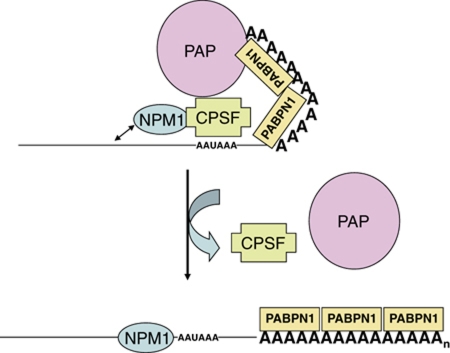
During polyadenylation, the multi-functional protein nucleophosmin (NPM1) is deposited onto all cellular mRNAs analysed to date. Premature termination of poly(A) tail synthesis in the presence of cordycepin abrogates deposition of the protein onto the mRNA, indicating natural termination of poly(A) addition is required for NPM1 binding. NPM1 appears to be a bona fide member of the complex involved in 3' end processing as it is associated with the AAUAAA-binding CPSF factor and can be co-immunoprecipitated with other polyadenylation factors. Furthermore, reduction in the levels of NPM1 results in hyperadenylation of mRNAs, consistent with alterations in poly(A) tail chain termination. Finally, knockdown of NPM1 results in retention of poly(A)(+) RNAs in the cell nucleus, indicating that NPM1 influences mRNA export. Collectively, these data suggest that NPM1 has an important role in poly(A) tail length determination and may help network 3' end processing with other aspects of nuclear mRNA maturation.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
Assembly of an export-competent mRNP is needed for efficient release of the 3'-end processing complex after polyadenylation.Mol Cell Biol. 2009 Oct;29(19):5327-38. doi: 10.1128/MCB.00468-09. Epub 2009 Jul 27. Mol Cell Biol. 2009. PMID: 19635808 Free PMC article.
-
Biophysical characterizations of the recognition of the AAUAAA polyadenylation signal.RNA. 2019 Dec;25(12):1673-1680. doi: 10.1261/rna.070870.119. Epub 2019 Aug 28. RNA. 2019. PMID: 31462423 Free PMC article.
-
Nucleophosmin is selectively deposited on mRNA during polyadenylation.Nat Struct Mol Biol. 2006 May;13(5):429-35. doi: 10.1038/nsmb1080. Epub 2006 Apr 9. Nat Struct Mol Biol. 2006. PMID: 16604083 Free PMC article.
-
Birth of a poly(A) tail: mechanisms and control of mRNA polyadenylation.FEBS Open Bio. 2023 Jul;13(7):1140-1153. doi: 10.1002/2211-5463.13528. Epub 2022 Dec 7. FEBS Open Bio. 2023. PMID: 36416579 Free PMC article. Review.
-
Implications of polyadenylation in health and disease.Nucleus. 2014;5(6):508-19. doi: 10.4161/nucl.36360. Epub 2014 Oct 31. Nucleus. 2014. PMID: 25484187 Free PMC article. Review.
Cited by
-
Adenoviral protein V promotes a process of viral assembly through nucleophosmin 1.Virology. 2012 Oct 25;432(2):283-95. doi: 10.1016/j.virol.2012.05.028. Epub 2012 Jun 18. Virology. 2012. PMID: 22717133 Free PMC article.
-
Chromosomal passenger complex hydrodynamics suggests chaperoning of the inactive state by nucleoplasmin/nucleophosmin.Mol Biol Cell. 2017 Jun 1;28(11):1444-1456. doi: 10.1091/mbc.E16-12-0860. Epub 2017 Apr 12. Mol Biol Cell. 2017. PMID: 28404751 Free PMC article.
-
Determinants and implications of mRNA poly(A) tail size--does this protein make my tail look big?Semin Cell Dev Biol. 2014 Oct;34:24-32. doi: 10.1016/j.semcdb.2014.05.018. Epub 2014 Jun 5. Semin Cell Dev Biol. 2014. PMID: 24910447 Free PMC article. Review.
-
An intrabody specific for the nucleophosmin carboxy-terminal mutant and fused to a nuclear localization sequence binds its antigen but fails to relocate it in the nucleus.Biotechnol Rep (Amst). 2014 May 27;3:27-33. doi: 10.1016/j.btre.2014.05.008. eCollection 2014 Sep. Biotechnol Rep (Amst). 2014. PMID: 28626645 Free PMC article.
-
Inhibition of polyadenylation reduces inflammatory gene induction.RNA. 2012 Dec;18(12):2236-50. doi: 10.1261/rna.032391.112. Epub 2012 Nov 1. RNA. 2012. PMID: 23118416 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
