

Behavioral deficit, oxidative stress, and mitochondrial dysfunction precede tau pathology in P301S transgenic mice
- PMID: 21825035
- PMCID: PMC3205832
- DOI: 10.1096/fj.11-186650
Behavioral deficit, oxidative stress, and mitochondrial dysfunction precede tau pathology in P301S transgenic mice
Abstract
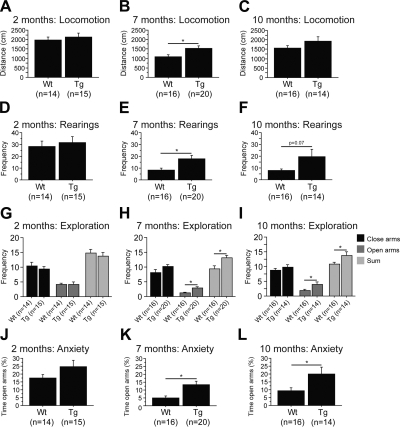
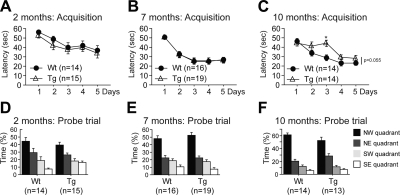
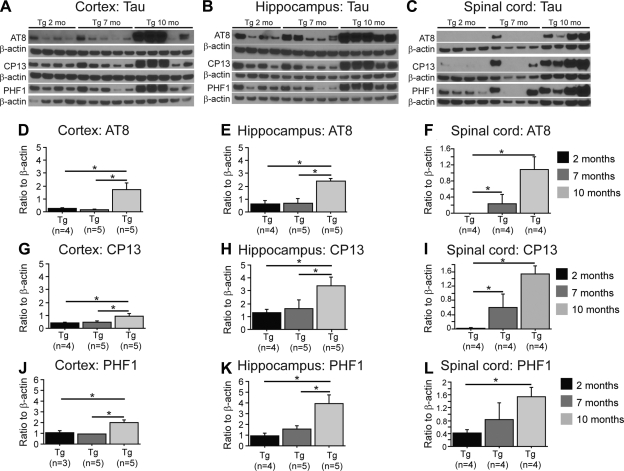
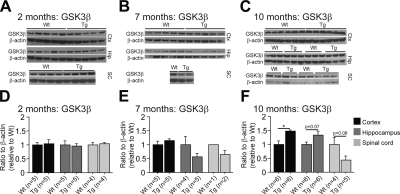
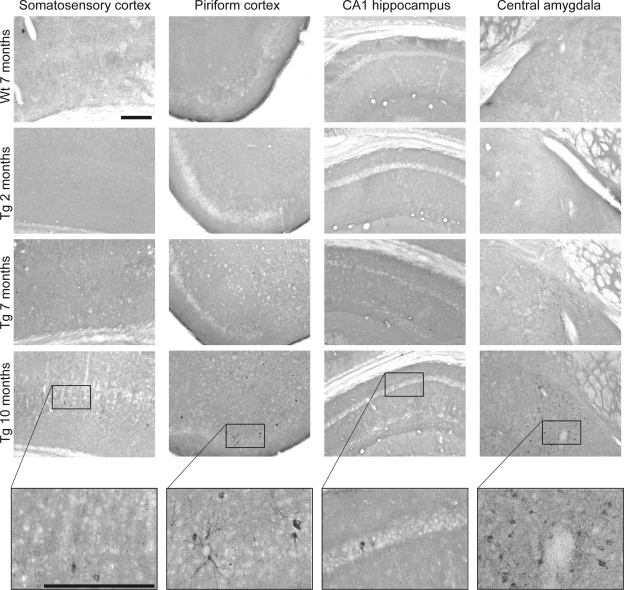
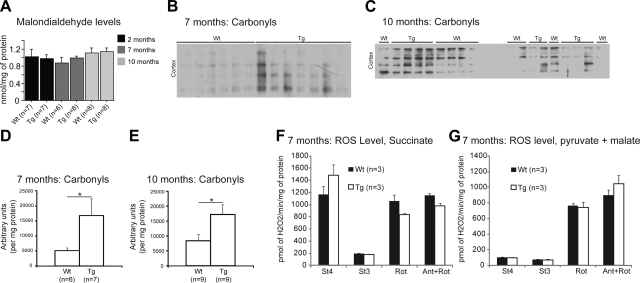
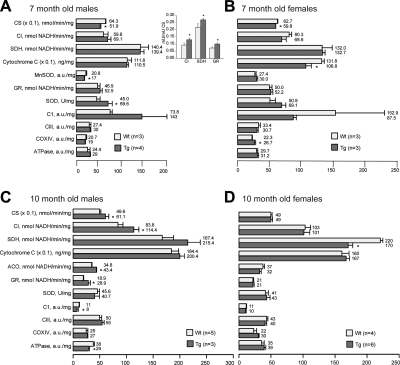
Abnormal tau accumulation can lead to the development of neurodegenerative diseases. P301S mice overexpress the human tau mutated gene, resulting in tau hyperphosphorylation and tangle formation. Mice also develop synaptic deficits and microglial activation prior to any neurodegeneration and tangles. Oxidative stress can also affect tauopathy. We studied the role of oxidative stress in relationship to behavioral abnormalities and disease progression in P301S mice at 2, 7, and 10 mo of age. At 7 mo of age, P301S mice had behavioral abnormalities, such as hyperactivity and disinhibition. At the same age, we observed increased carbonyls in P301S mitochondria (∼215 and 55% increase, males/females), and deregulation in the activity and content of mitochondrial enzymes involved in reactive oxygen species formation and energy metabolism, such as citrate synthase (∼19 and ∼5% decrease, males/females), MnSOD (∼16% decrease, males only), cytochrome C (∼19% decrease, females only), and cytochrome C oxidase (∼20% increase, females only). These changes in mitochondria proteome appeared before tau hyperphosphorylation and tangle formation, which were observed at 10 mo and were associated with GSK3β activation. At that age, mitochondria proteome deregulation became more apparent in male P301S mitochondria. The data strongly suggest that oxidative stress and mitochondrial abnormalities appear prior to tau pathology.
Figures
References
-
- Lee V. M., Goedert M., Trojanowski J. Q. (2001) Neurodegenerative tauopathies. Annu. Rev. Neurosci. 24, 1121–1159 - PubMed
-
- Yoshiyama Y., Lee V. M., Trojanowski J. Q. (2001) Frontotemporal dementia and tauopathy. Curr. Neurol. Neurosci. Rep. 1, 413–421 - PubMed
-
- Gasparini L., Terni B., Spillantini M. G. (2007) Frontotemporal dementia with tau pathology. Neurodegener. Dis. 4, 236–253 - PubMed
-
- Goedert M., Jakes R. (2005) Mutations causing neurodegenerative tauopathies. Biochim. Biophys. Acta 1739, 240–250 - PubMed
-
- Ingram E. M., Spillantini M. G. (2002) Tau gene mutations: dissecting the pathogenesis of FTDP-17. Trends Mol. Med. 8, 555–562 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
