

Impaired parasite attachment as fitness cost of metronidazole resistance in Giardia lamblia
- PMID: 21825286
- PMCID: PMC3186953
- DOI: 10.1128/AAC.00384-11
Impaired parasite attachment as fitness cost of metronidazole resistance in Giardia lamblia
Abstract
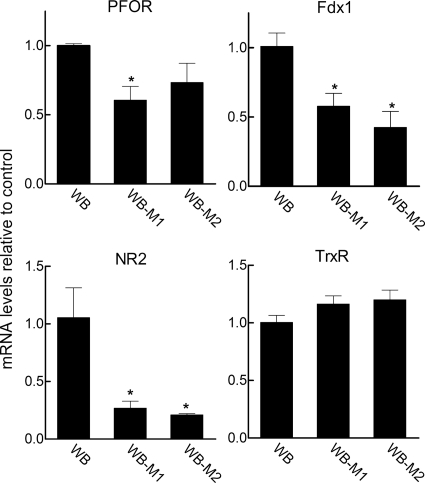
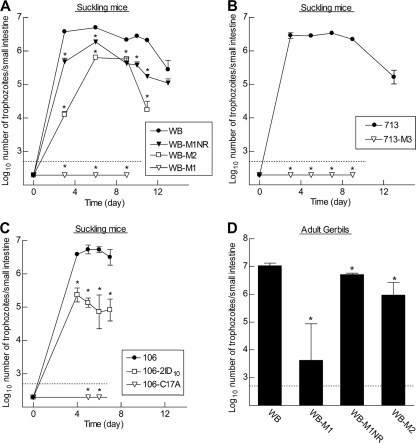
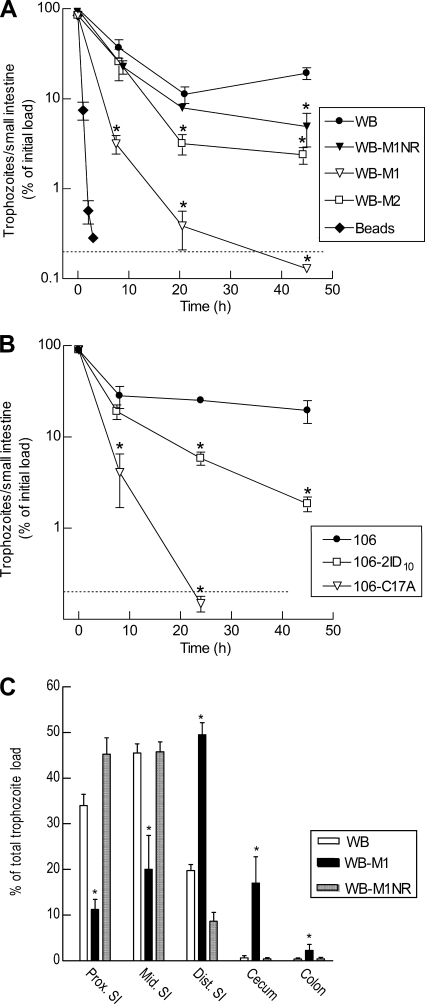
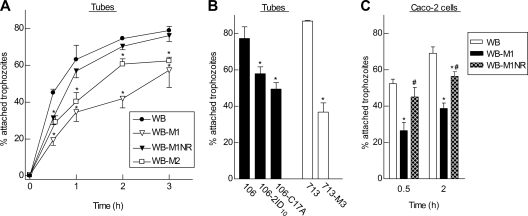
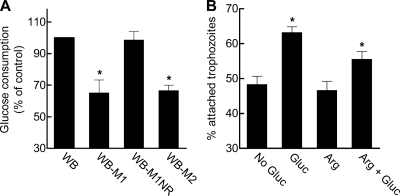
Infections with the diarrheagenic protozoan pathogen Giardia lamblia are most commonly treated with metronidazole (Mz). Treatment failures with Mz occur in 10 to 20% of cases and Mz resistance develops in the laboratory, yet clinically, Mz-resistant (Mz(r)) G. lamblia has rarely been isolated from patients. To understand why clinical Mz(r) isolates are rare, we questioned whether Mz resistance entails fitness costs to the parasite. Our studies employed several newly generated and established isogenic Mz(r) cell lines with stable, high-level resistance to Mz and significant cross-resistance to tinidazole, nitazoxanide, and furazolidone. Oral infection of suckling mice revealed that three of five Mz(r) cell lines could not establish infection, while two Mz(r) cell lines infected pups, albeit with reduced efficiencies. Failure to colonize resulted from a diminished capacity of the parasite to attach to the intestinal mucosa in vivo and to epithelial cells and plastic surfaces in vitro. The attachment defect was related to impaired glucose metabolism, since the noninfectious Mz(r) lines consumed less glucose, and glucose promoted ATP-independent parasite attachment in the parental lines. Thus, resistance of Giardia to Mz is accompanied by a glucose metabolism-related attachment defect that can interfere with colonization of the host. Because glucose-metabolizing pathways are important for activation of the prodrug Mz, it follows that a fitness trade-off exists between diminished Mz activation and reduced infectivity, which may explain the observed paucity of clinical Mz(r) isolates of Giardia. However, the data also caution that some forms of Mz resistance do not markedly interfere with in vivo infectivity.
Figures
References
-
- Ankarklev J., Jerlstrom-Hultqvist J., Ringqvist E., Troell K., Svard S. G. 2010. Behind the smile: cell biology and disease mechanisms of Giardia species. Nat. Rev. Microbiol. 8:413–422 - PubMed
-
- Arguello-Garcia R., Cruz-Soto M., Romero-Montoya L., Ortega-Pierres G. 2009. In vitro resistance to 5-nitroimidazoles and benzimidazoles in Giardia duodenalis: variability and variation in gene expression. Infect. Genet. Evol. 9:1057–1064 - PubMed
-
- Bloch T., Davis T. E., Jr., Schwenk G. R., Jr 1987. Giardia lamblia in peritoneal fluid. Acta Cytol. 31:783–784 - PubMed
-
- Boreham P. F., Phillips R. E., Shepherd R. W. 1986. The activity of drugs against Giardia intestinalis in neonatal mice. J. Antimicrob. Chemother. 18:393–398 - PubMed
-
- Boreham P. F., Phillips R. E., Shepherd R. W. 1988. Altered uptake of metronidazole in vitro by stocks of Giardia intestinalis with different drug sensitivities. Trans. R. Soc. Trop. Med. Hyg. 82:104–106 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
