

Metabolic deficiences revealed in the biotechnologically important model bacterium Escherichia coli BL21(DE3)
- PMID: 21826210
- PMCID: PMC3149613
- DOI: 10.1371/journal.pone.0022830
Metabolic deficiences revealed in the biotechnologically important model bacterium Escherichia coli BL21(DE3)
Abstract
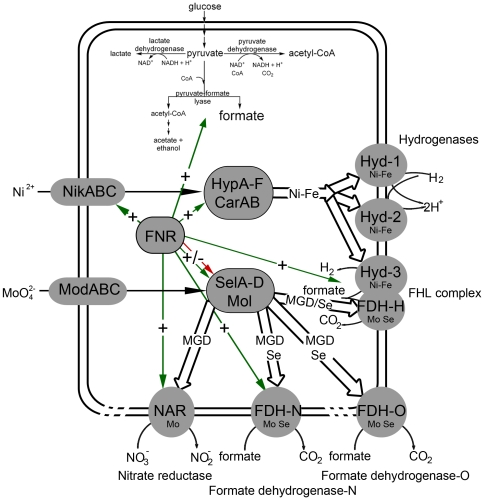
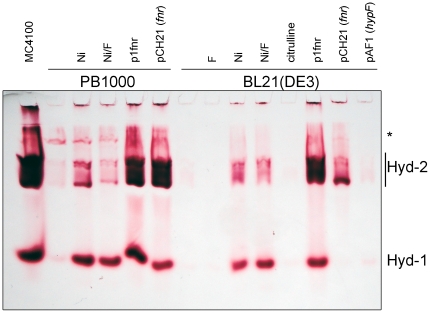
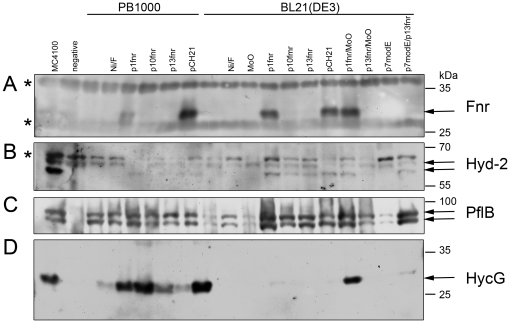
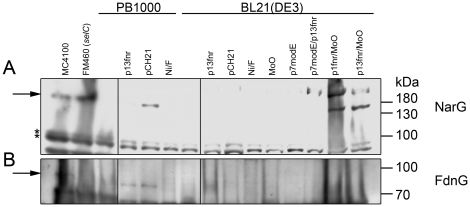
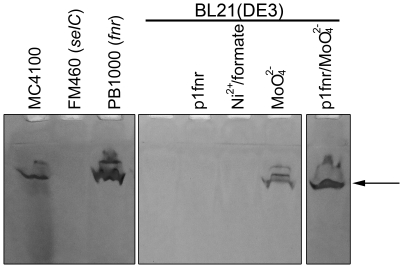
The Escherichia coli B strain BL21(DE3) has had a profound impact on biotechnology through its use in the production of recombinant proteins. Little is understood, however, regarding the physiology of this important E. coli strain. We show here that BL21(DE3) totally lacks activity of the four [NiFe]-hydrogenases, the three molybdenum- and selenium-containing formate dehydrogenases and molybdenum-dependent nitrate reductase. Nevertheless, all of the structural genes necessary for the synthesis of the respective anaerobic metalloenzymes are present in the genome. However, the genes encoding the high-affinity molybdate transport system and the molybdenum-responsive transcriptional regulator ModE are absent from the genome. Moreover, BL21(DE3) has a nonsense mutation in the gene encoding the global oxygen-responsive transcriptional regulator FNR. The activities of the two hydrogen-oxidizing hydrogenases, therefore, could be restored to BL21(DE3) by supplementing the growth medium with high concentrations of Ni²⁺ (Ni²⁺-transport is FNR-dependent) or by introducing a wild-type copy of the fnr gene. Only combined addition of plasmid-encoded fnr and high concentrations of MoO₄²⁻ ions could restore hydrogen production to BL21(DE3); however, to only 25-30% of a K-12 wildtype. We could show that limited hydrogen production from the enzyme complex responsible for formate-dependent hydrogen evolution was due solely to reduced activity of the formate dehydrogenase (FDH-H), not the hydrogenase component. The activity of the FNR-dependent formate dehydrogenase, FDH-N, could not be restored, even when the fnr gene and MoO₄²⁻ were supplied; however, nitrate reductase activity could be recovered by combined addition of MoO₄²⁻ and the fnr gene. This suggested that a further component specific for biosynthesis or activity of formate dehydrogenases H and N was missing. Re-introduction of the gene encoding ModE could only partially restore the activities of both enzymes. Taken together these results demonstrate that BL21(DE3) has major defects in anaerobic metabolism, metal ion transport and metalloprotein biosynthesis.
Conflict of interest statement
Figures
Similar articles
-
Activation of formate hydrogen-lyase via expression of uptake [NiFe]-hydrogenase in Escherichia coli BL21(DE3).Microb Cell Fact. 2015 Sep 22;14:151. doi: 10.1186/s12934-015-0343-0. Microb Cell Fact. 2015. PMID: 26395073 Free PMC article.
-
Differential effects of isc operon mutations on the biosynthesis and activity of key anaerobic metalloenzymes in Escherichia coli.Microbiology (Reading). 2017 Jun;163(6):878-890. doi: 10.1099/mic.0.000481. Epub 2017 Jun 26. Microbiology (Reading). 2017. PMID: 28640740
-
The respiratory molybdo-selenoprotein formate dehydrogenases of Escherichia coli have hydrogen: benzyl viologen oxidoreductase activity.BMC Microbiol. 2011 Aug 1;11:173. doi: 10.1186/1471-2180-11-173. BMC Microbiol. 2011. PMID: 21806784 Free PMC article.
-
The hydrogenases and formate dehydrogenases of Escherichia coli.Antonie Van Leeuwenhoek. 1994;66(1-3):57-88. doi: 10.1007/BF00871633. Antonie Van Leeuwenhoek. 1994. PMID: 7747941 Review.
-
Multiple and reversible hydrogenases for hydrogen production by Escherichia coli: dependence on fermentation substrate, pH and the F(0)F(1)-ATPase.Crit Rev Biochem Mol Biol. 2012 May-Jun;47(3):236-49. doi: 10.3109/10409238.2012.655375. Epub 2012 Feb 7. Crit Rev Biochem Mol Biol. 2012. PMID: 22313414 Review.
Cited by
-
Trace element associated reduction of norleucine and norvaline accumulation during oxygen limitation in a recombinant Escherichia coli fermentation.Microb Cell Fact. 2013 Nov 21;12:116. doi: 10.1186/1475-2859-12-116. Microb Cell Fact. 2013. PMID: 24261588 Free PMC article.
-
Genetic and Biochemical Analysis of the Azotobacter vinelandii Molybdenum Storage Protein.Front Microbiol. 2019 Mar 21;10:579. doi: 10.3389/fmicb.2019.00579. eCollection 2019. Front Microbiol. 2019. PMID: 30984129 Free PMC article.
-
An innovative cloning platform enables large-scale production and maturation of an oxygen-tolerant [NiFe]-hydrogenase from Cupriavidus necator in Escherichia coli.PLoS One. 2013 Jul 5;8(7):e68812. doi: 10.1371/journal.pone.0068812. Print 2013. PLoS One. 2013. PMID: 23861944 Free PMC article.
-
Iron restriction induces preferential down-regulation of H(2)-consuming over H(2)-evolving reactions during fermentative growth of Escherichia coli.BMC Microbiol. 2011 Aug 31;11:196. doi: 10.1186/1471-2180-11-196. BMC Microbiol. 2011. PMID: 21880124 Free PMC article.
-
Short-term differential adaptation to anaerobic stress via genomic mutations by Escherichia coli strains K-12 and B lacking alcohol dehydrogenase.Front Microbiol. 2014 Sep 9;5:476. doi: 10.3389/fmicb.2014.00476. eCollection 2014. Front Microbiol. 2014. PMID: 25250024 Free PMC article.
References
-
- Sawers R, Clark D, Böck A. Fermentative Pyruvate and Acetyl-Coenzyme A Metabolism. 2004. EcoSal - Escherichia coli and Salmonella: cellular and molecular biology. - PubMed
-
- Spiro S, Guest JR. Adaptive responses to oxygen limitation in Escherichia coli. Trends Biochem Sci. 1991;16:310–314. - PubMed
-
- Schindelin H, Kisker C, Rajagopalan KV. Molybdopterin from molybdenum and tungsten enzymes. Adv Protein Chem. 2001;58:47–94. - PubMed
-
- Forzi L, Sawers RG. Maturation of [NiFe]-hydrogenases in Escherichia coli. Biometals. 2007;20:565–578. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
