

Type 1 interferon induction of natural killer cell gamma interferon production for defense during lymphocytic choriomeningitis virus infection
- PMID: 21828218
- PMCID: PMC3150756
- DOI: 10.1128/mBio.00169-11
Type 1 interferon induction of natural killer cell gamma interferon production for defense during lymphocytic choriomeningitis virus infection
Abstract
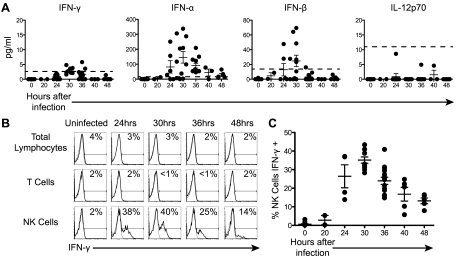
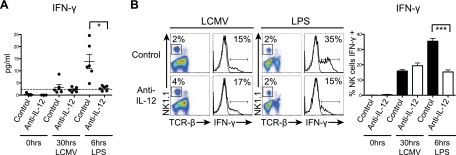
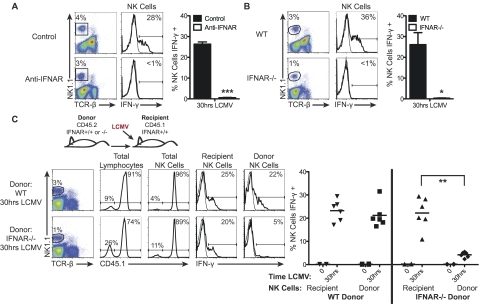
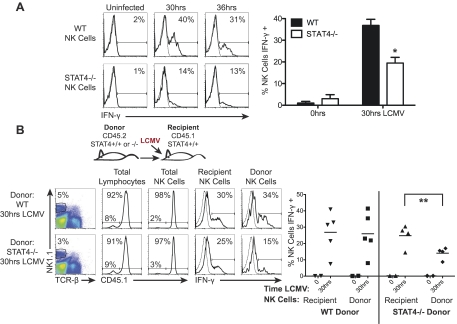
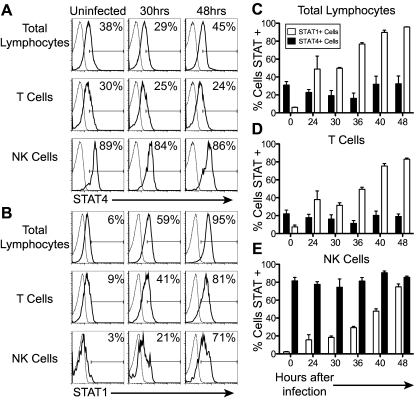
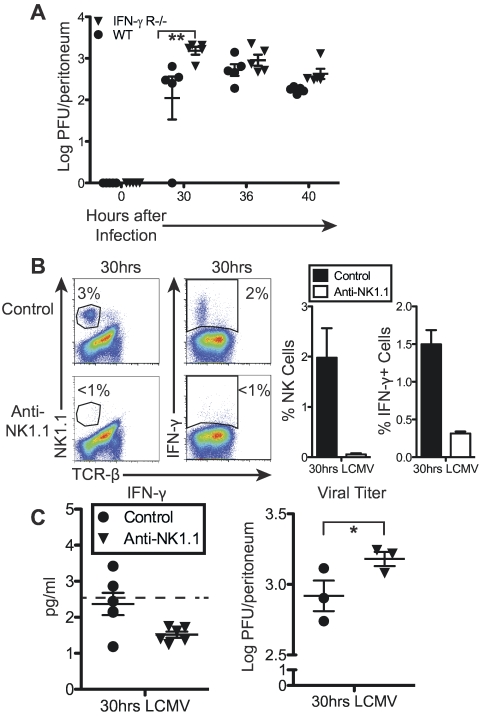
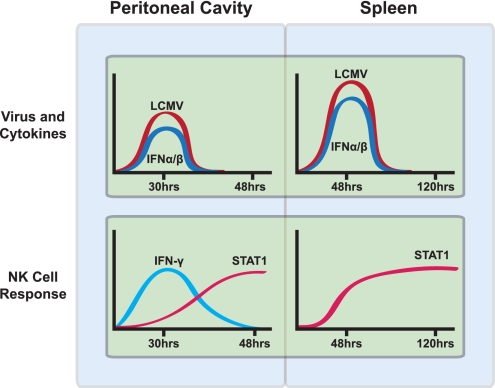
Natural killer (NK) cells are equipped to innately produce the cytokine gamma interferon (IFN-γ) in part because they basally express high levels of the signal transducer and activator of transcription 4 (STAT4). Type 1 interferons (IFNs) have the potential to activate STAT4 and promote IFN-γ expression, but concurrent induction of elevated STAT1 negatively regulates access to the pathway. As a consequence, it has been difficult to detect type 1 IFN stimulation of NK cell IFN-γ during viral infections in the presence of STAT1 and to understand the evolutionary advantage for maintaining the pathway. The studies reported here evaluated NK cell responses following infections with lymphocytic choriomeningitis virus (LCMV) in the compartment handling the earliest events after infection, the peritoneal cavity. The production of type 1 IFNs, both IFN-α and IFN-β, was shown to be early and of short duration, peaking at 30 h after challenge. NK cell IFN-γ expression was detected with overlapping kinetics and required activating signals delivered through type 1 IFN receptors and STAT4. It took place under conditions of high STAT4 levels but preceded elevated STAT1 expression in NK cells. The IFN-γ response reduced viral burdens. Interestingly, increases in STAT1 were delayed in NK cells compared to other peritoneal exudate cell (PEC) populations. Taken together, the studies demonstrate a novel mechanism for stimulating IFN-γ production and elucidate a biological role for type 1 IFN access to STAT4 in NK cells.
Importance: Pathways regulating the complex and sometimes paradoxical effects of cytokines are poorly understood. Accumulating evidence indicates that the biological consequences of type 1 interferon (IFN) exposure are shaped by modifying the concentrations of particular STATs to change access to the different signaling molecules. The results of the experiments presented conclusively demonstrate that NK cell IFN-γ can be induced through type 1 IFN and STAT4 at the first site of infection during a period with high STAT4 but prior to induction of elevated STAT1 in the cells. The response mediates a role in viral defense. Thus, a very early pathway to and source of IFN-γ in evolving immune responses to infections are identified by this work. The information obtained helps resolve long-standing controversies and advances the understanding of mechanisms regulating key type 1 IFN functions, in different cells and compartments and at different times of infection, for accessing biologically important functions.
Figures
References
-
- Biron CA, Nguyen KB, Pien GC, Cousens LP, Salazar-Mather TP. 1999. Natural killer cells in antiviral defense: function and regulation by innate cytokines. Annu. Rev. Immunol. 17:189–220 - PubMed
-
- Orange JS, Biron CA. 1996. Characterization of early IL-12, IFN-alphabeta, and TNF effects on antiviral state and NK cell responses during murine cytomegalovirus infection. J. Immunol. 156:4746–4756 - PubMed
-
- Nguyen KB, et al. 2002. Coordinated and distinct roles for IFN-alpha beta, IL-12, and IL-15 regulation of NK cell responses to viral infection. J. Immunol. 169:4279–4287 - PubMed
-
- García-Sastre A, Biron CA. 2006. Type 1 interferons and the virus-host relationship: a lesson in detente. Science 312:879–882 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
