

Sustained activation of the HER1-ERK1/2-RSK signaling pathway controls myoepithelial cell fate in human mammary tissue
- PMID: 21828273
- PMCID: PMC3182019
- DOI: 10.1101/gad.2025611
Sustained activation of the HER1-ERK1/2-RSK signaling pathway controls myoepithelial cell fate in human mammary tissue
Abstract
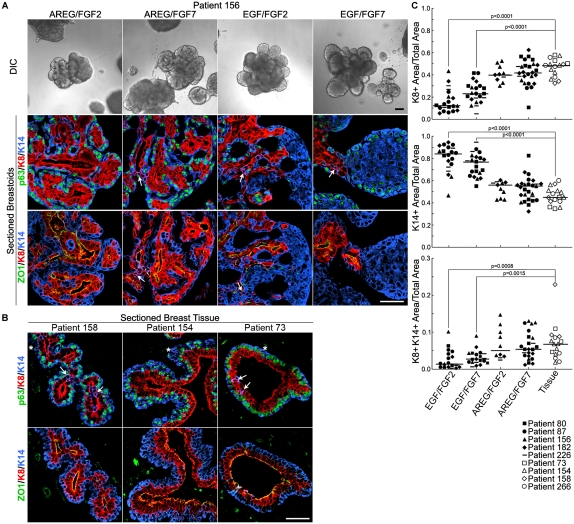
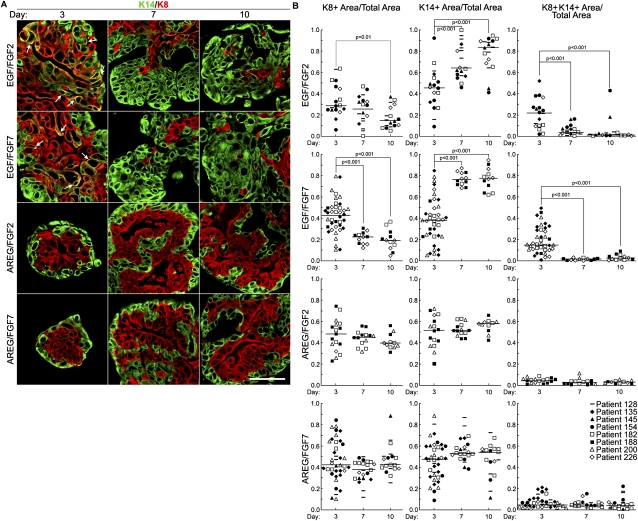
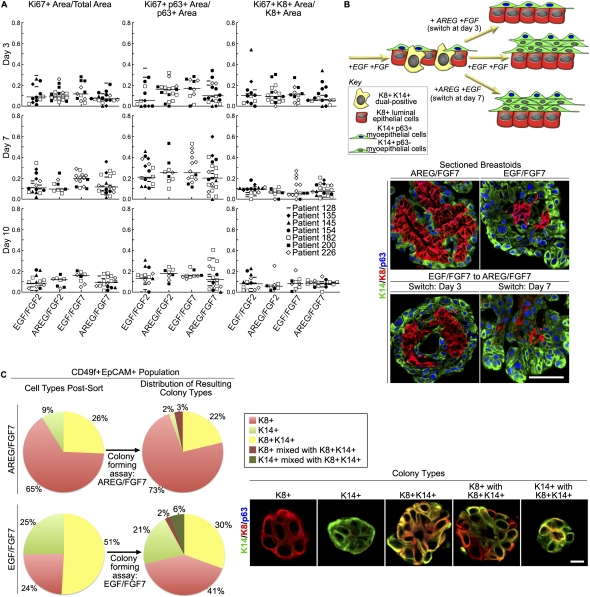
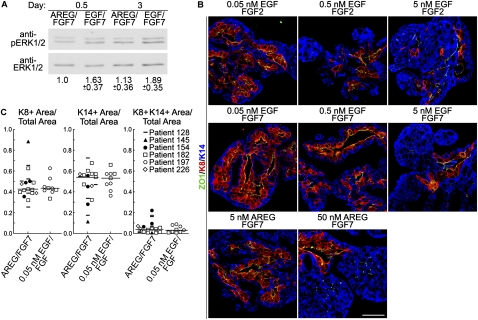
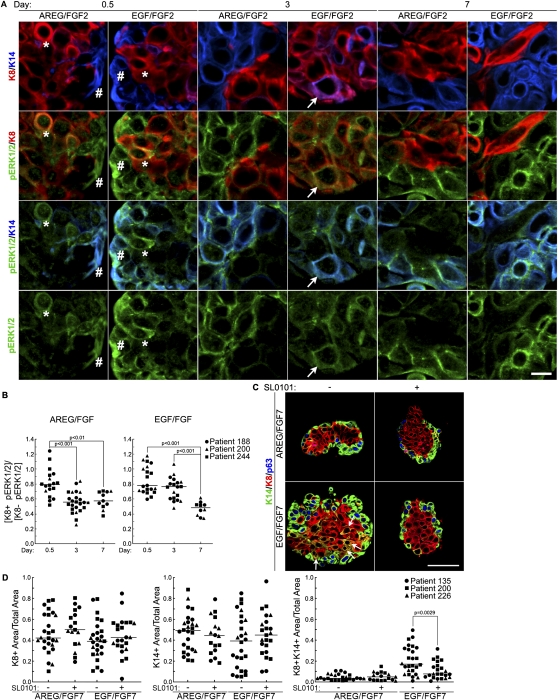
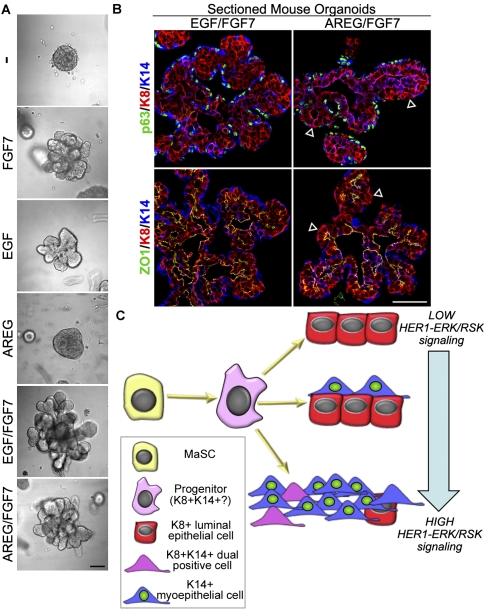
Human mammary glands arise from multipotent progenitor cells, which likely respond both to cell-autonomous and to extrinsic cues. However, the identity of these cues and how they might act remain unclear. We analyzed HER1 ligand effects on mammary morphogenesis using a three-dimensional organoid model generated from human breast tissue that recapitulates both qualitatively and quantitatively the normal ductal network in situ. Strikingly, different HER1 ligands generate distinct patterns of cell fate. Epidermal growth factor (EGF) causes a massive expansion of the myoepithelial lineage. Amphiregulin, in contrast, enables normal ductal development. These differences cannot be ascribed to preferential apoptosis or proliferation of differentiated cell populations, but are dependent on HER1 signal intensity. Inhibition of the extracellular signal-regulated kinase 1/2 (ERK1/2) effector RSK prevents the EGF-induced myoepithelial expansion. Notably, mouse mammary organoids are much less responsive to HER1 ligands. Little is known about the myoepithelial lineage or about growth factor effects on mammary progenitor differentiation, and our studies provide an important window into human mammary development that reveals unexpected differences from the mouse model.
Figures
References
-
- Andrechek ER, White D, Muller WJ 2005. Targeted disruption of ErbB2/Neu in the mammary epithelium results in impaired ductal outgrowth. Oncogene 24: 932–937 - PubMed
-
- Asselin-Labat ML, Vaillant F, Sheridan JM, Pal B, Wu D, Simpson ER, Yasuda H, Smyth GK, Martin TJ, Lindeman GJ, et al. 2010. Control of mammary stem cell function by steroid hormone signalling. Nature 465: 798–802 - PubMed
-
- Barsky SH, Karlin NJ 2006. Mechanisms of disease: breast tumor pathogenesis and the role of the myoepithelial cell. Nat Clin Pract Oncol 3: 138–151 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous