

Plastid uridine salvage activity is required for photoassimilate allocation and partitioning in Arabidopsis
- PMID: 21828290
- PMCID: PMC3180806
- DOI: 10.1105/tpc.111.085829
Plastid uridine salvage activity is required for photoassimilate allocation and partitioning in Arabidopsis
Abstract
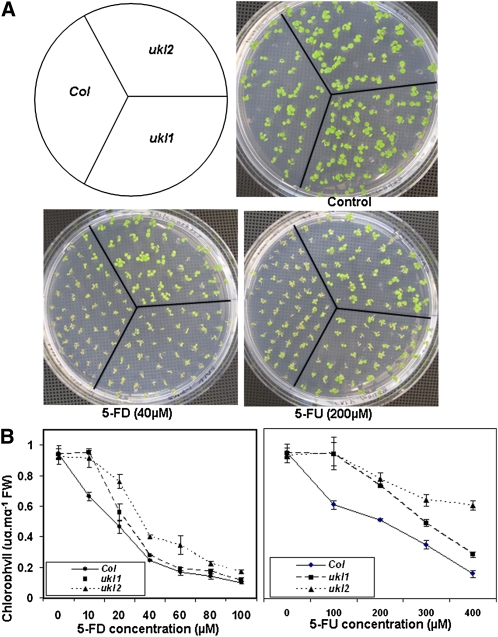
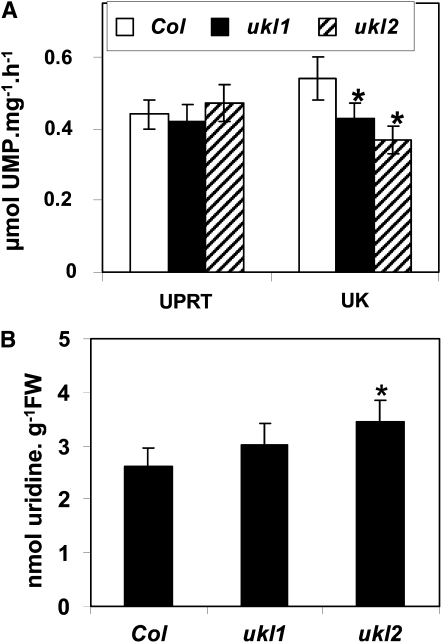
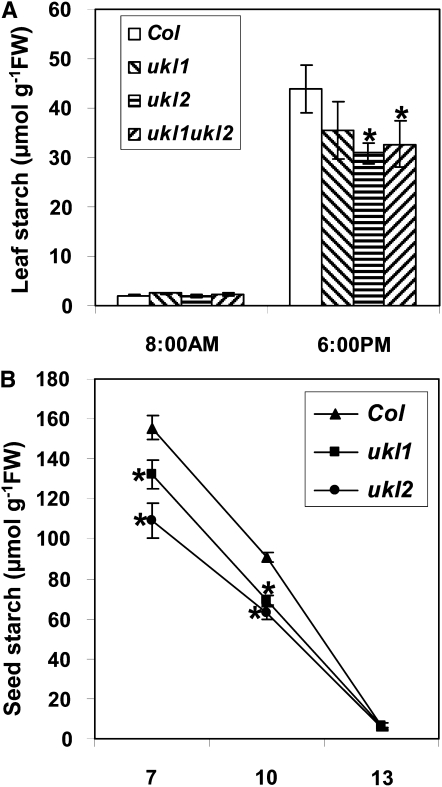
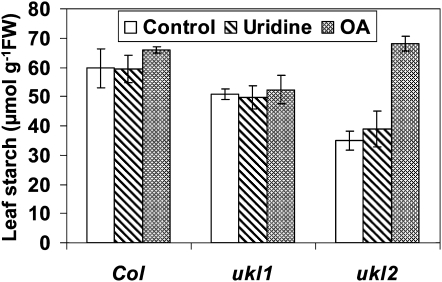
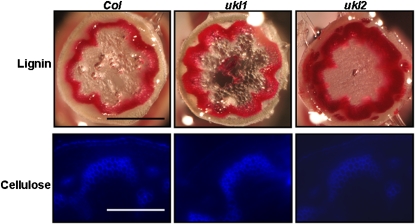
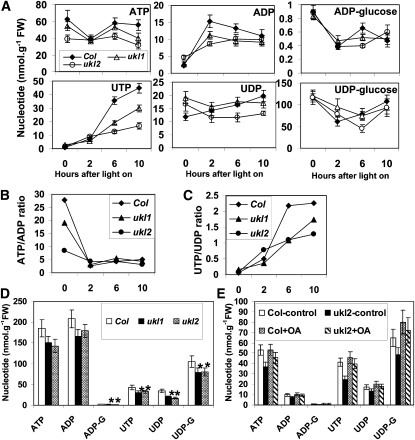
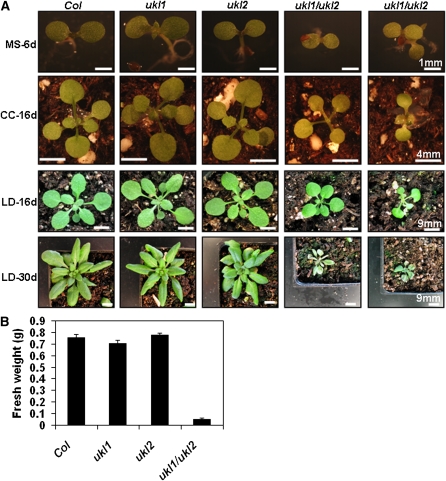
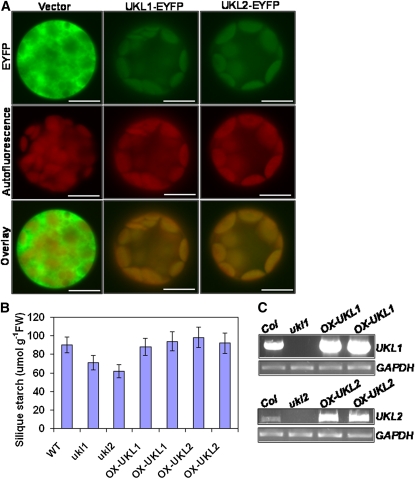
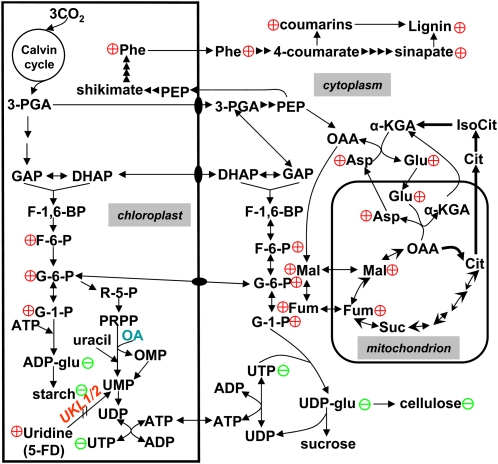
Nucleotides are synthesized from de novo and salvage pathways. To characterize the uridine salvage pathway, two genes, UKL1 and UKL2, that tentatively encode uridine kinase (UK) and uracil phosphoribosyltransferase (UPRT) bifunctional enzymes were studied in Arabidopsis thaliana. T-DNA insertions in UKL1 and UKL2 reduced transcript expression and increased plant tolerance to toxic analogs 5-fluorouridine and 5-fluorouracil. Enzyme activity assays using purified recombinant proteins indicated that UKL1 and UKL2 have UK but not UPRT activity. Subcellular localization using a C-terminal enhanced yellow fluorescent protein fusion indicated that UKL1 and UKL2 localize to plastids. The ukl2 mutant shows reduced transient leaf starch during the day. External application of orotate rescued this phenotype in ukl2, indicating pyrimidine pools are limiting for starch synthesis in ukl2. Intermediates for lignin synthesis were upregulated, and there was increased lignin and reduced cellulose content in the ukl2 mutant. Levels of ATP, ADP, ADP-glucose, UTP, UDP, and UDP-glucose were altered in a light-dependent manner. Seed composition of the ukl1 and ukl2 mutants included lower oil and higher protein compared with the wild type. Unlike single gene mutants, the ukl1 ukl2 double mutant has severe developmental defects and reduced biomass accumulation, indicating these enzymes catalyze redundant reactions. These findings point to crucial roles played by uridine salvage for photoassimilate allocation and partitioning.
Figures
References
-
- Anterola A.M., Jeon J.H., Davin L.B., Lewis N.G. (2002). Transcriptional control of monolignol biosynthesis in Pinus taeda: Factors affecting monolignol ratios and carbon allocation in phenylpropanoid metabolism. J. Biol. Chem. 277: 18272–18280 - PubMed
-
- Ashihara H. (1977). Changes in the activities of the de novo and salvage pathways of pyrimidine nucleotide biosynthesis during germination of black gram (Phaseolus mungo) seeds. Z. Pflanzenphysiol. 81: 199–211
-
- Boerjan W., Ralph J., Baucher M. (2003). Lignin biosynthesis. Annu. Rev. Plant Biol. 54: 519–546 - PubMed
-
- Bolwell G.P., Butt V.S., Davies D.R., Zimmerlin A. (1995). The origin of the oxidative burst in plants. Free Radic. Res. 23: 517–532 - PubMed
-
- Broeckling C.D., Reddy I.R., Duran A.L., Zhao X.C., Sumner L.W. (2006). MET-IDEA: Data extraction tool for mass spectrometry-based metabolomics. Anal. Chem. 78: 4334–4341 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
