

Cortical state and attention
- PMID: 21829219
- PMCID: PMC3324821
- DOI: 10.1038/nrn3084
Cortical state and attention
Abstract
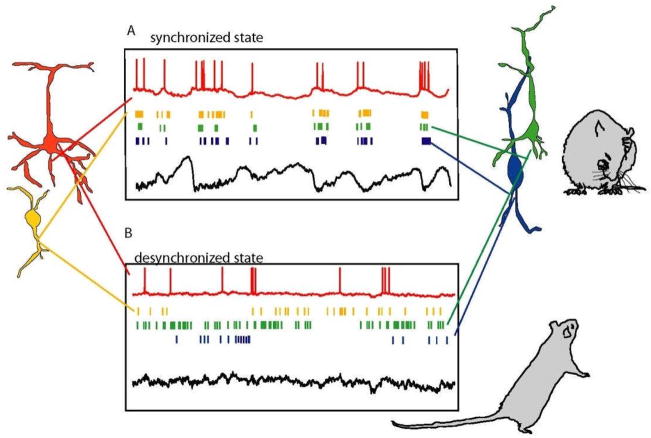
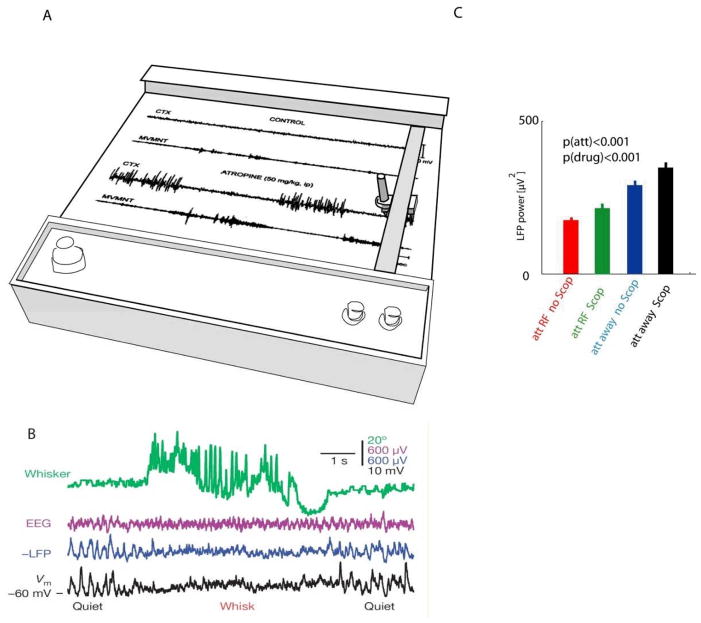
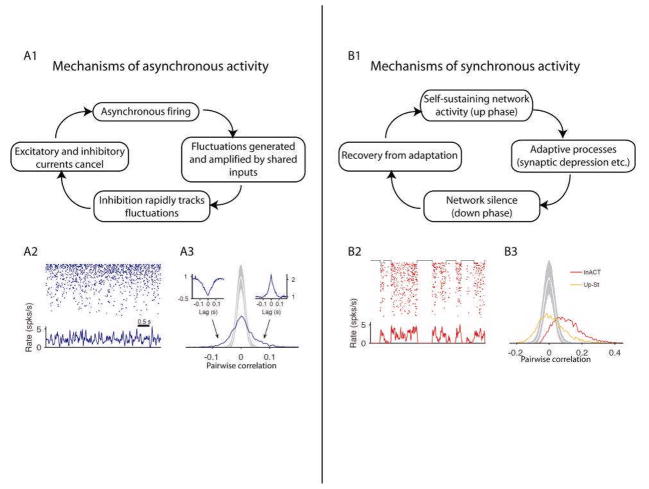
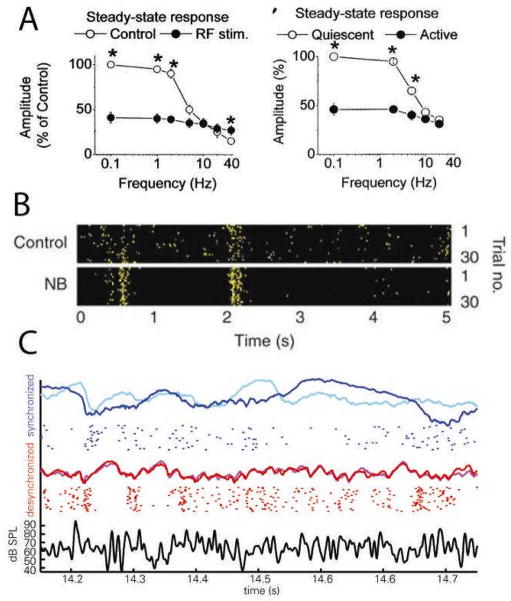
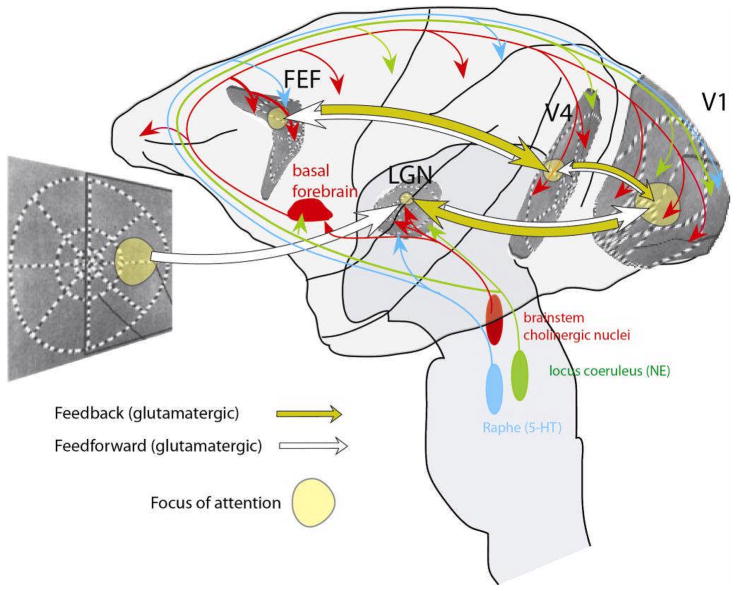
The brain continuously adapts its processing machinery to behavioural demands. To achieve this, it rapidly modulates the operating mode of cortical circuits, controlling the way that information is transformed and routed. This article will focus on two experimental approaches by which the control of cortical information processing has been investigated: the study of state-dependent cortical processing in rodents and attention in the primate visual system. Both processes involve a modulation of low-frequency activity fluctuations and spiking correlation, and are mediated by common receptor systems. We suggest that selective attention involves processes that are similar to state change, and that operate at a local columnar level to enhance the representation of otherwise non-salient features while suppressing internally generated activity patterns.
Figures
References
-
- Steriade M, McCarley RW. Brainstem control of wakefulness and sleep. Plenum Press; New York: 2005.
-
- Greenberg DS, Houweling AR, Kerr JN. Population imaging of ongoing neuronal activity in the visual cortex of awake rats. Nat Neurosci. 2008;11:749–51. - PubMed
-
- Crochet S, Petersen CC. Correlating whisker behavior with membrane potential in barrel cortex of awake mice. Nat Neurosci. 2006;9:608–10. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
