

An epigenetic switch involving overlapping fur and DNA methylation optimizes expression of a type VI secretion gene cluster
- PMID: 21829382
- PMCID: PMC3145626
- DOI: 10.1371/journal.pgen.1002205
An epigenetic switch involving overlapping fur and DNA methylation optimizes expression of a type VI secretion gene cluster
Abstract
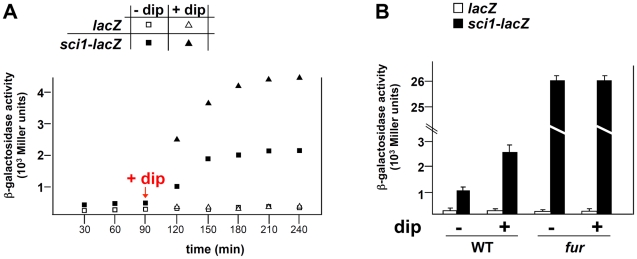
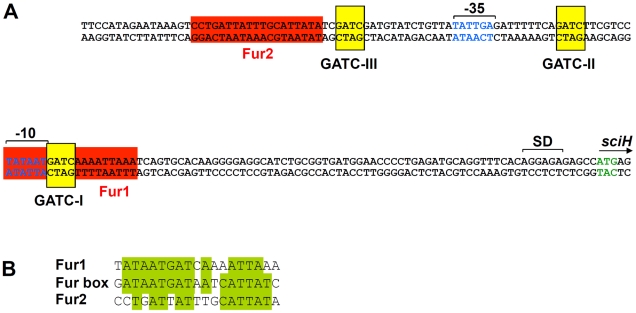
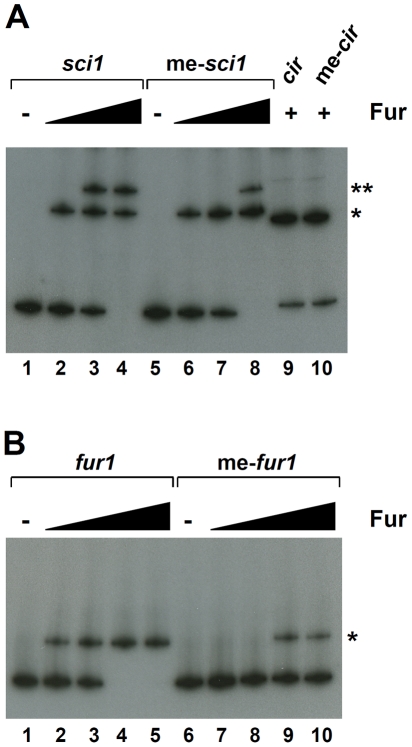
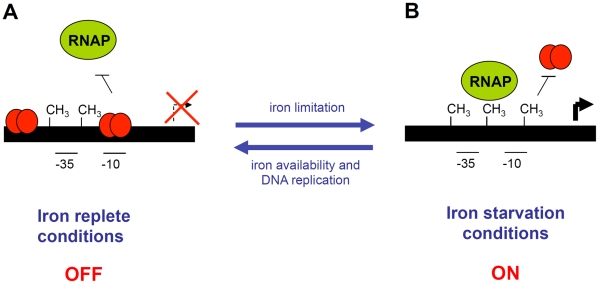
Type VI secretion systems (T6SS) are macromolecular machines of the cell envelope of Gram-negative bacteria responsible for bacterial killing and/or virulence towards different host cells. Here, we characterized the regulatory mechanism underlying expression of the enteroagregative Escherichia coli sci1 T6SS gene cluster. We identified Fur as the main regulator of the sci1 cluster. A detailed analysis of the promoter region showed the presence of three GATC motifs, which are target of the DNA adenine methylase Dam. Using a combination of reporter fusion, gel shift, and in vivo and in vitro Dam methylation assays, we dissected the regulatory role of Fur and Dam-dependent methylation. We showed that the sci1 gene cluster expression is under the control of an epigenetic switch depending on methylation: fur binding prevents methylation of a GATC motif, whereas methylation at this specific site decreases the affinity of Fur for its binding box. A model is proposed in which the sci1 promoter is regulated by iron availability, adenine methylation, and DNA replication.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



Similar articles
-
Fur-Dam Regulatory Interplay at an Internal Promoter of the Enteroaggregative Escherichia coli Type VI Secretion sci1 Gene Cluster.J Bacteriol. 2020 Apr 27;202(10):e00075-20. doi: 10.1128/JB.00075-20. Print 2020 Apr 27. J Bacteriol. 2020. PMID: 32152218 Free PMC article.
-
The mechanism by which DNA adenine methylase and PapI activate the pap epigenetic switch.Mol Cell. 2003 Oct;12(4):947-57. doi: 10.1016/s1097-2765(03)00383-6. Mol Cell. 2003. PMID: 14580345
-
Pyrimidine regulation of the Escherichia coli and Salmonella typhimurium carAB operons: CarP and integration host factor (IHF) modulate the methylation status of a GATC site present in the control region.J Mol Biol. 1995 Jul 21;250(4):383-91. doi: 10.1006/jmbi.1995.0384. J Mol Biol. 1995. PMID: 7616563
-
Members of the Fur protein family regulate iron and zinc transport in E. coli and characteristics of the Fur-regulated fhuF protein.J Mol Microbiol Biotechnol. 2002 May;4(3):217-22. J Mol Microbiol Biotechnol. 2002. PMID: 11931550 Review.
-
The intricate workings of a bacterial epigenetic switch.Adv Exp Med Biol. 2004;547:83-9. doi: 10.1007/978-1-4419-8861-4_7. Adv Exp Med Biol. 2004. PMID: 15230094 Review.
Cited by
-
Structure and regulation of the type VI secretion system.Annu Rev Microbiol. 2012;66:453-72. doi: 10.1146/annurev-micro-121809-151619. Epub 2012 Jun 28. Annu Rev Microbiol. 2012. PMID: 22746332 Free PMC article. Review.
-
Bidirectional contraction of a type six secretion system.Nat Commun. 2019 Apr 5;10(1):1565. doi: 10.1038/s41467-019-09603-1. Nat Commun. 2019. PMID: 30952865 Free PMC article.
-
Regulation of bistability in the std fimbrial operon of Salmonella enterica by DNA adenine methylation and transcription factors HdfR, StdE and StdF.Nucleic Acids Res. 2019 Sep 5;47(15):7929-7941. doi: 10.1093/nar/gkz530. Nucleic Acids Res. 2019. PMID: 31216025 Free PMC article.
-
DNA Adenine Methyltransferase (Dam) Overexpression Impairs Photorhabdus luminescens Motility and Virulence.Front Microbiol. 2017 Sep 1;8:1671. doi: 10.3389/fmicb.2017.01671. eCollection 2017. Front Microbiol. 2017. PMID: 28919886 Free PMC article.
-
The Type VI Secretion System in Escherichia coli and Related Species.EcoSal Plus. 2016 May;7(1):10.1128/ecosalplus.ESP-0009-2015. doi: 10.1128/ecosalplus.ESP-0009-2015. EcoSal Plus. 2016. PMID: 27223818 Free PMC article. Review.
References
-
- Bingle LE, Bailey CM, Pallen MJ. Type VI secretion: a beginner's guide. Curr Opin Microbiol. 2008;11:3–8. - PubMed
-
- Filloux A, Hachani A, Bleves S. The bacterial type VI secretion machine: yet another player for protein transport across membranes. Microbiology. 2008;154:1570–83. - PubMed
-
- Aschtgen MS, Gavioli M, Dessen A, Lloubès R, Cascales E. The SciZ protein anchors the enteroaggregative Escherichia coli Type VI secretion system to the cell wall. Mol Microbiol. 2010;75:886–99. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
