

The X-linked tumor suppressor TSPX interacts and promotes degradation of the hepatitis B viral protein HBx via the proteasome pathway
- PMID: 21829568
- PMCID: PMC3146538
- DOI: 10.1371/journal.pone.0022979
The X-linked tumor suppressor TSPX interacts and promotes degradation of the hepatitis B viral protein HBx via the proteasome pathway
Abstract
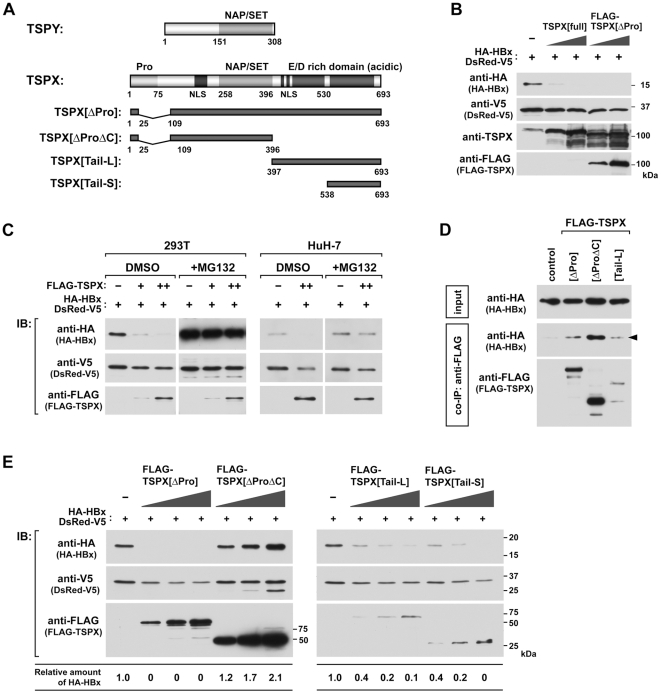
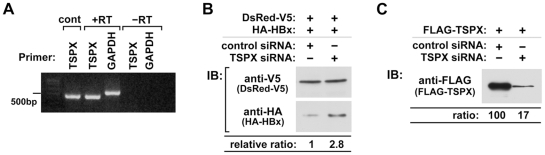
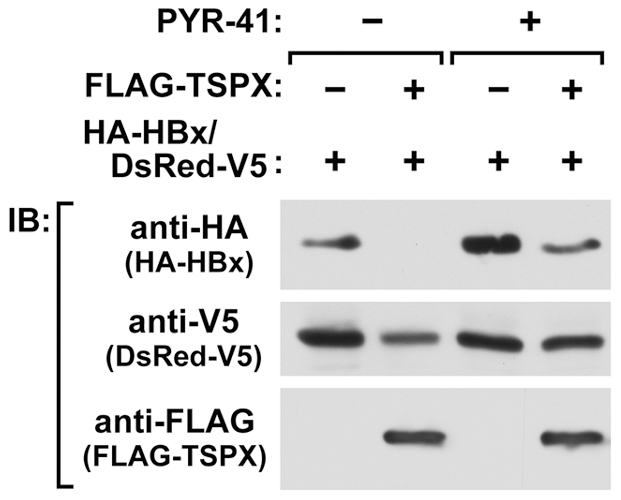
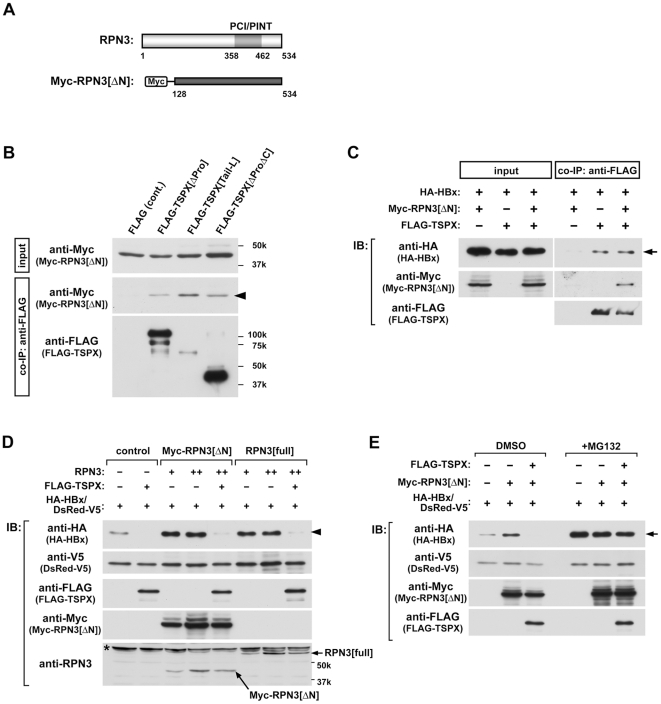
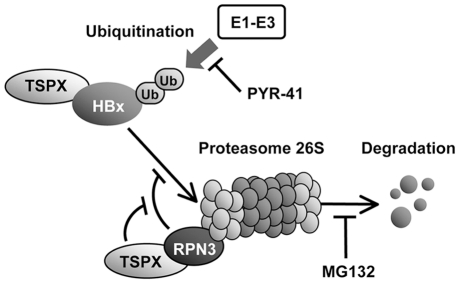
Hepatitis B virus (HBV) infection is a major risk for hepatocellular carcinoma (HCC), and it is a serious global health problem with two billion people exposed to it worldwide. HBx, an essential factor for viral replication and a putative oncoprotein encoded by the HBV genome, has been shown to promote oncogenic properties at multiple sites in HBV-infected liver cells. The expression level of HBx closely associates with the development and progression of HCC, therefore the mechanism(s) regulating the stability of HBx is important in oncogenesis of HBV-infected cells. We demonstrate that the X-linked tumor suppressor TSPX enhances the degradation of HBx through the ubiquitin-proteasome pathway. TSPX interacts with both HBx and a proteasome 19S lid subunit RPN3 via its C-terminal acidic tail. Most importantly, over-expression of RPN3 protects HBx from, and hence acts as a negative regulator for, proteasome-dependent degradation. TSPX abrogates the RPN3-depedent stabilization of HBx, suggesting that TSPX and RPN3 act competitively in regulation of HBx stability. Since mutation and/or epigenetic repression of X-located tumor suppressor gene(s) could significantly predispose males to human cancers, our data suggest that TSPX-induced HBx degradation could play key role(s) in hepatocarcinogenesis among HBV-infected HCC patients.
Conflict of interest statement
Figures
Similar articles
-
Hepatitis B Virus X Protein Is Stabilized by the Deubiquitinating Enzyme VCPIP1 in a Ubiquitin-Independent Manner by Recruiting the 26S Proteasome Subunit PSMC3.J Virol. 2022 Jul 13;96(13):e0061122. doi: 10.1128/jvi.00611-22. Epub 2022 Jun 13. J Virol. 2022. PMID: 35695579 Free PMC article.
-
E3 ubiquitin ligase Siah-1 facilitates poly-ubiquitylation and proteasomal degradation of the hepatitis B viral X protein.FEBS Lett. 2011 Oct 3;585(19):2943-50. doi: 10.1016/j.febslet.2011.08.015. Epub 2011 Aug 26. FEBS Lett. 2011. PMID: 21878328
-
Hepatitis B virus X protein promotes hepatocellular carcinoma transformation through interleukin-6 activation of microRNA-21 expression.Eur J Cancer. 2014 Oct;50(15):2560-9. doi: 10.1016/j.ejca.2014.07.008. Epub 2014 Jul 30. Eur J Cancer. 2014. PMID: 25087183
-
Modulation of apoptotic signaling by the hepatitis B virus X protein.Viruses. 2012 Nov 8;4(11):2945-72. doi: 10.3390/v4112945. Viruses. 2012. PMID: 23202511 Free PMC article. Review.
-
Impact of hepatitis B virus X protein on the DNA damage response during hepatocarcinogenesis.Med Mol Morphol. 2009 Sep;42(3):138-42. doi: 10.1007/s00795-009-0457-8. Epub 2009 Sep 26. Med Mol Morphol. 2009. PMID: 19784739 Review.
Cited by
-
The interaction of hepatitis B virus with the ubiquitin proteasome system in viral replication and associated pathogenesis.Virol J. 2019 May 30;16(1):73. doi: 10.1186/s12985-019-1183-z. Virol J. 2019. PMID: 31146743 Free PMC article. Review.
-
The Y-located proto-oncogene TSPY exacerbates and its X-homologue TSPX inhibits transactivation functions of androgen receptor and its constitutively active variants.Hum Mol Genet. 2017 Mar 1;26(5):901-912. doi: 10.1093/hmg/ddx005. Hum Mol Genet. 2017. PMID: 28169398 Free PMC article.
-
Battle of the sexes: contrasting roles of testis-specific protein Y-encoded (TSPY) and TSPX in human oncogenesis.Asian J Androl. 2019 May-Jun;21(3):260-269. doi: 10.4103/aja.aja_43_18. Asian J Androl. 2019. PMID: 29974883 Free PMC article. Review.
-
Sexual dimorphism in cancer.Nat Rev Cancer. 2016 May;16(5):330-9. doi: 10.1038/nrc.2016.30. Epub 2016 Apr 15. Nat Rev Cancer. 2016. PMID: 27079803 Review.
-
The X-linked tumor suppressor TSPX downregulates cancer-drivers/oncogenes in prostate cancer in a C-terminal acidic domain dependent manner.Oncotarget. 2019 Feb 19;10(15):1491-1506. doi: 10.18632/oncotarget.26673. eCollection 2019 Feb 19. Oncotarget. 2019. PMID: 30863497 Free PMC article.
References
-
- Park YJ, Luger K. Structure and function of nucleosome assembly proteins. Biochem Cell Biol. 2006;84:549–558. - PubMed
-
- Ozbun LL, Martinez A, Jakowlew SB. Differentially expressed nucleolar TGF-beta1 target (DENTT) shows tissue-specific nuclear and cytoplasmic localization and increases TGF-beta1-responsive transcription in primates. Biochim Biophys Acta. 2005;1728:163–180. - PubMed
-
- Scully RE. Gonadoblastoma. A review of 74 cases. Cancer. 1970;25:1340–1356. - PubMed
-
- Page DC. Hypothesis: a Y-chromosomal gene causes gonadoblastoma in dysgenetic gonads. Development. 1987;101(Suppl):151–155. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
