

Protein accumulation in leaves and roots associated with improved drought tolerance in creeping bentgrass expressing an ipt gene for cytokinin synthesis
- PMID: 21831843
- PMCID: PMC3223035
- DOI: 10.1093/jxb/err166
Protein accumulation in leaves and roots associated with improved drought tolerance in creeping bentgrass expressing an ipt gene for cytokinin synthesis
Abstract
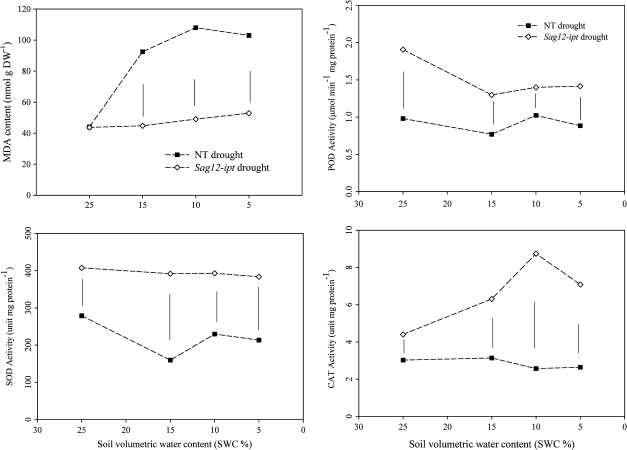
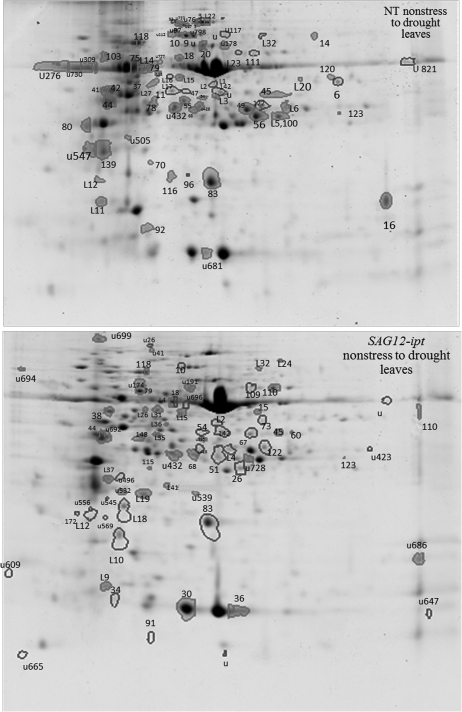
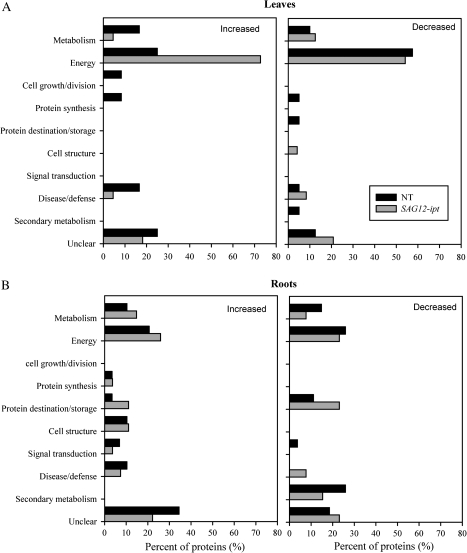
Cytokinins (CKs) may be involved in the regulation of plant adaptation to drought stress. The objectives of the study were to identify proteomic changes in leaves and roots in relation to improved drought tolerance in transgenic creeping bentgrass (Agrostis stolonifera) containing a senescence-activated promoter (SAG12) and the isopentyl transferase (ipt) transgene that increases endogenous CK content. Leaves of SAG12-ipt bentgrass exhibited less severe senescence under water stress, as demonstrated by maintaining lower electrolyte leakage and lipid peroxidation, and higher photochemical efficiency (F(v)/F(m)), compared with the null transformant (NT) plants. SAG12-ipt plants had higher root/shoot ratios and lower lipid peroxidation in leaves under water stress than the NT plants. The suppression of drought-induced leaf senescence and root dieback in the transgenic plants was associated with the maintenance of greater antioxidant enzyme activities (superoxide dismutase, peroxidase, and catalase). The SAG12-ipt and NT plants exhibited differential protein expression patterns under well-watered and drought conditions in both leaves and roots. Under equivalent leaf water deficit (47% relative water content), SAG12-ipt plants maintained higher abundance of proteins involved in (i) energy production within both photosynthesis and respiration [ribulose 1,5-bisphosphate carboxylase (RuBisCO) and glyceraldehyde phosphate dehydrogenase (GAPDH)]; (ii) amino acid synthesis (methionine and glutamine); (iii) protein synthesis and destination [chloroplastic elongation factor (EF-Tu) and protein disulphide isomerases (PDIs)]; and (iv) antioxidant defence system (catalase and peroxidase) than the NT plants. These results suggest that increased endogenous CKs under drought stress may directly or indirectly regulate protein abundance and enzymatic activities involved in the above-mentioned metabolic processes, thereby enhancing plant drought tolerance.
Figures




References
-
- Baek D, Jin Y, Jeong JC, et al. Suppression of reactive oxygen species by glyceraldehyde-3-phosphate dehydrogenase. Phytochemistry. 2008;69:333–338. - PubMed
-
- Bagniewska-Zadworna A. The root microtubule cytoskeleton and cell cycle analysis through desiccation of Brassica napus seedlings. Protoplasma. 2008;233:177–185. - PubMed
-
- Barrs HD, Weatherley PE. A re-examination of the relative turgidity techniques for estimating water deficits in leaves. Australian Journal of Biological Sciences. 1962;15:413–428.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
