

Spinal locomotor inputs to individually identified reticulospinal neurons in the lamprey
- PMID: 21832033
- PMCID: PMC3214102
- DOI: 10.1152/jn.01100.2010
Spinal locomotor inputs to individually identified reticulospinal neurons in the lamprey
Abstract
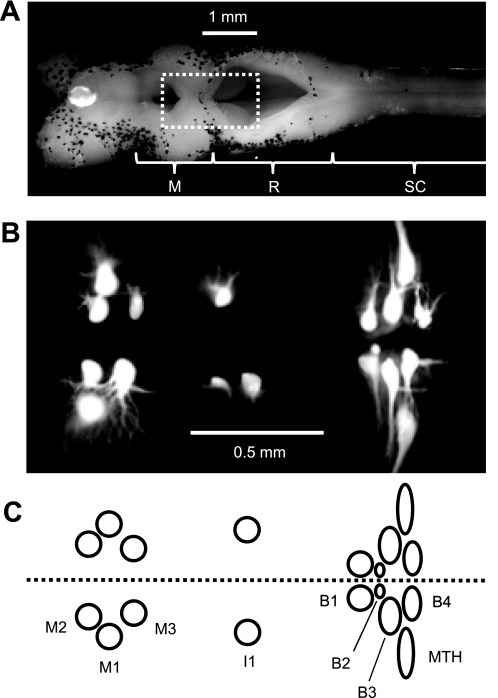
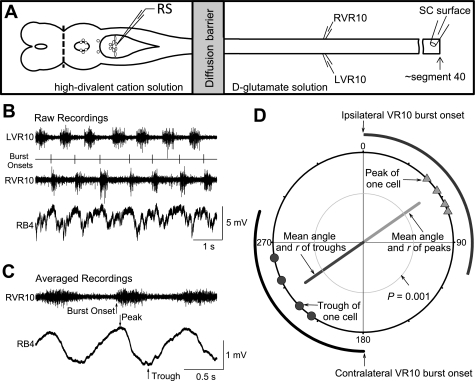
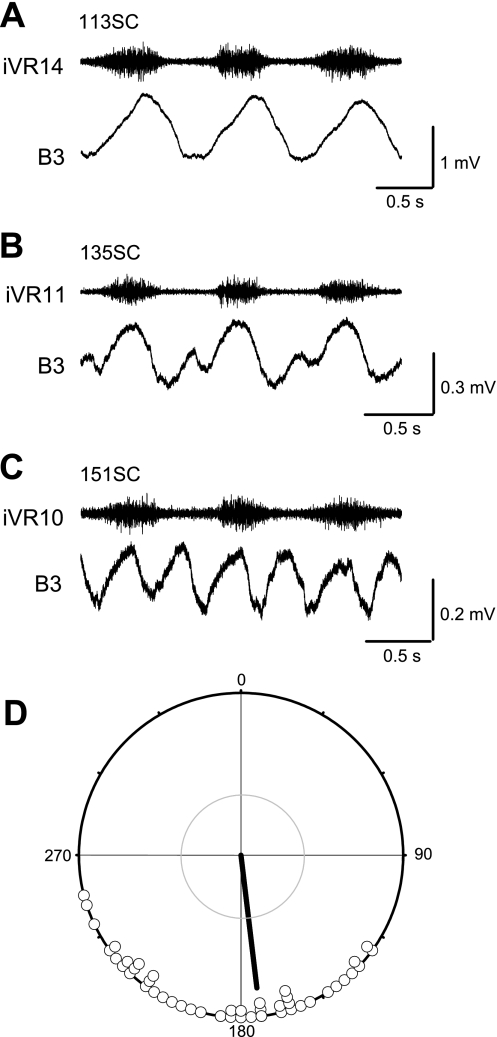
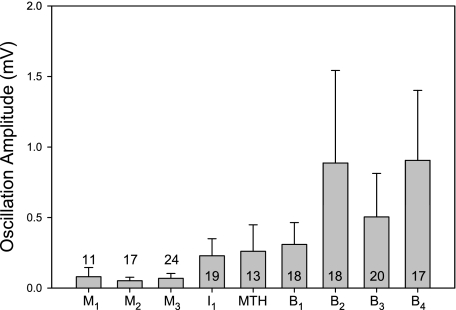
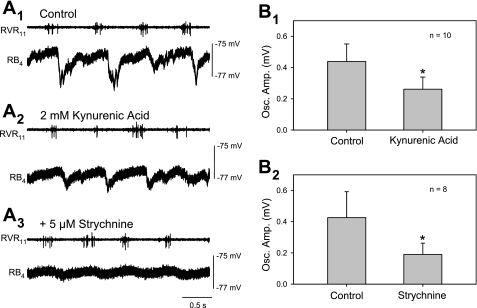
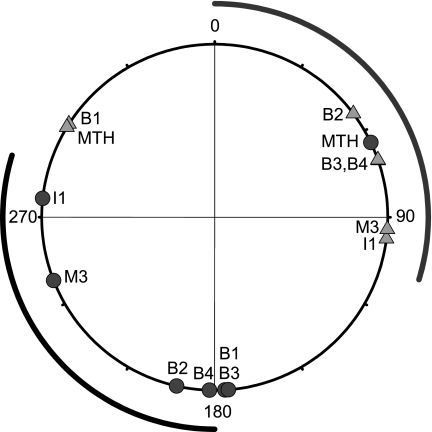
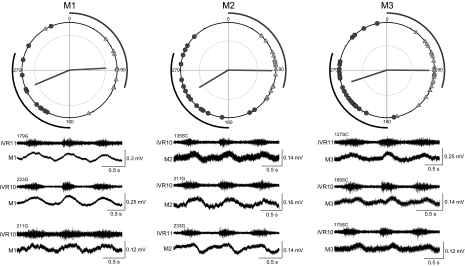
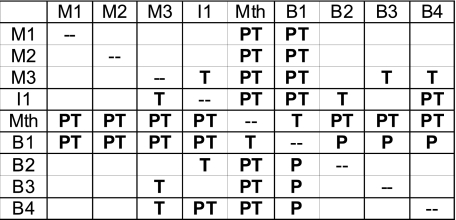
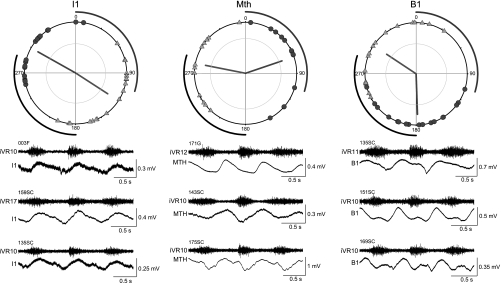
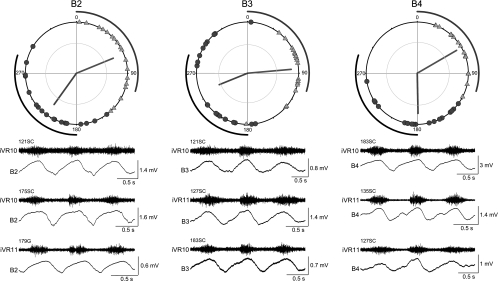
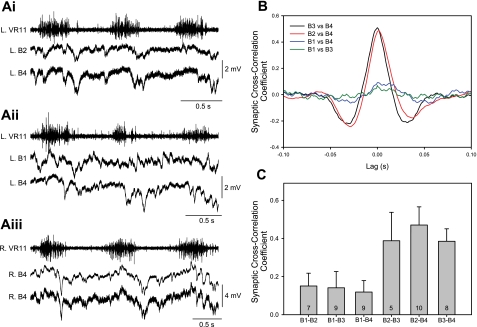
Locomotor feedback signals from the spinal cord to descending brain stem neurons were examined in the lamprey using the uniquely identifiable reticulospinal neurons, the Müller and Mauthner cells. The same identified reticulospinal neurons were recorded in several preparations, under reduced conditions, to address whether an identified reticulospinal neuron shows similar locomotor-related oscillation timing from animal to animal and whether these timing signals can differ significantly from other identified reticulospinal neurons. Intracellular recordings of membrane potential in identified neurons were made in an isolated brain stem-spinal cord preparation with a high-divalent cation solution on the brain stem to suppress indirect neural pathways and with D-glutamate perfusion to the spinal cord to induce fictive swimming. Under these conditions, the identified reticulospinal neurons show significant clustering of the timings of the peaks and troughs of their locomotor-related oscillations. Whereas most identified neurons oscillated in phase with locomotor bursting in ipsilateral ventral roots of the rostral spinal cord, the B1 Müller cell, which has an ipsilateral descending axon, and the Mauthner cell, which has a contralateral descending axon, both had oscillation peaks that were out of phase with the ipsilateral ventral roots. The differences in oscillation timing appear to be due to differences in synaptic input sources as shown by cross-correlations of fast synaptic activity in pairs of Müller cells. Since the main source of the locomotor input under these experimental conditions is ascending neurons in the spinal cord, these experiments suggest that individual reticulospinal neurons can receive locomotor signals from different subsets of these ascending neurons. This result may indicate that the locomotor feedback signals from the spinal locomotor networks are matched in some way to the motor output functions of the individual reticulospinal neurons, which include command signals for turning and for compensatory movements.
Figures
Similar articles
-
Contributions of identifiable neurons and neuron classes to lamprey vertebrate neurobiology.Prog Neurobiol. 2001 Mar;63(4):441-66. doi: 10.1016/s0301-0082(00)00050-2. Prog Neurobiol. 2001. PMID: 11163686 Review.
-
Müller cells and other reticulospinal neurones are phasically active during fictive locomotion in the isolated nervous system of the lamprey.Neurosci Lett. 1986 Sep 12;69(3):239-43. doi: 10.1016/0304-3940(86)90486-6. Neurosci Lett. 1986. PMID: 3763053
-
Reticulospinal neurons receive direct spinobulbar inputs during locomotor activity in lamprey.J Neurophysiol. 2004 Sep;92(3):1384-90. doi: 10.1152/jn.00625.2003. J Neurophysiol. 2004. PMID: 15331645
-
Membrane potential oscillations in reticulospinal and spinobulbar neurons during locomotor activity.J Neurophysiol. 2005 Jul;94(1):273-81. doi: 10.1152/jn.00695.2004. Epub 2005 Mar 2. J Neurophysiol. 2005. PMID: 15744013
-
The spinobulbar system in lamprey.Brain Res Rev. 2008 Jan;57(1):37-45. doi: 10.1016/j.brainresrev.2007.07.010. Epub 2007 Aug 6. Brain Res Rev. 2008. PMID: 17716741 Free PMC article. Review.
Cited by
-
Cholinergic Modulation of Locomotor Circuits in Vertebrates.Int J Mol Sci. 2022 Sep 14;23(18):10738. doi: 10.3390/ijms231810738. Int J Mol Sci. 2022. PMID: 36142651 Free PMC article. Review.
-
Necessary, sufficient and permissive: a single locomotor command neuron important for intersegmental coordination.J Neurosci. 2012 Dec 5;32(49):17646-57. doi: 10.1523/JNEUROSCI.2249-12.2012. J Neurosci. 2012. PMID: 23223287 Free PMC article.
-
Reticulospinal Systems for Tuning Motor Commands.Front Neural Circuits. 2018 Apr 18;12:30. doi: 10.3389/fncir.2018.00030. eCollection 2018. Front Neural Circuits. 2018. PMID: 29720934 Free PMC article. Review.
-
Swimming rhythm generation in the caudal hindbrain of the lamprey.J Neurophysiol. 2018 May 1;119(5):1681-1692. doi: 10.1152/jn.00851.2017. Epub 2018 Jan 24. J Neurophysiol. 2018. PMID: 29364070 Free PMC article.
-
Tectal microcircuit generating visual selection commands on gaze-controlling neurons.Proc Natl Acad Sci U S A. 2015 Apr 14;112(15):E1956-65. doi: 10.1073/pnas.1504866112. Epub 2015 Mar 30. Proc Natl Acad Sci U S A. 2015. PMID: 25825743 Free PMC article.
References
-
- Arshavsky YI, Berkinblit MB, Fukson OI, Gelfand IM, Orlovsky GN. Recordings of neurons of the dorsal spinocerebellar tract during evoked locomotion. Brain Res 43: 272–275, 1972 - PubMed
-
- Arshavsky YI, Gelfand IM, Orlovsky GN, Pavlova GA. Messages conveyed by spinocerebellar pathways during scratching in the cat. I. Activity of neurons of the lateral reticular nucleus. Brain Res 151: 479–491, 1978a - PubMed
-
- Arshavsky YI, Gelfand IM, Orlovsky GN, Pavlova GA. Messages conveyed by spinocerebellar pathways during scratching in the cat. II. Activity of neurons of the ventral spinocerebellar tract. Brain Res 151:493–506, 1978b - PubMed
-
- Berry MS, Pentreath VW. Criteria for distinguishing between monosynaptic and polysynaptic transmission. Brain Res 105: 1–20, 1976 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources