

Sensory input regulates spatial and subtype-specific patterns of neuronal turnover in the adult olfactory bulb
- PMID: 21832189
- PMCID: PMC6623130
- DOI: 10.1523/JNEUROSCI.0614-11.2011
Sensory input regulates spatial and subtype-specific patterns of neuronal turnover in the adult olfactory bulb
Abstract
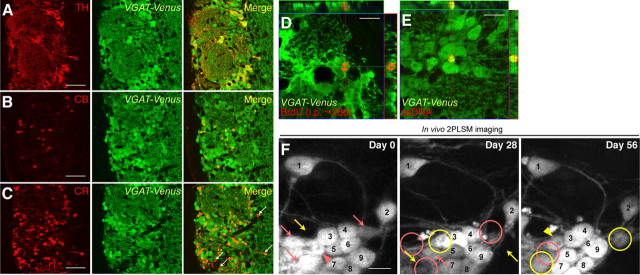
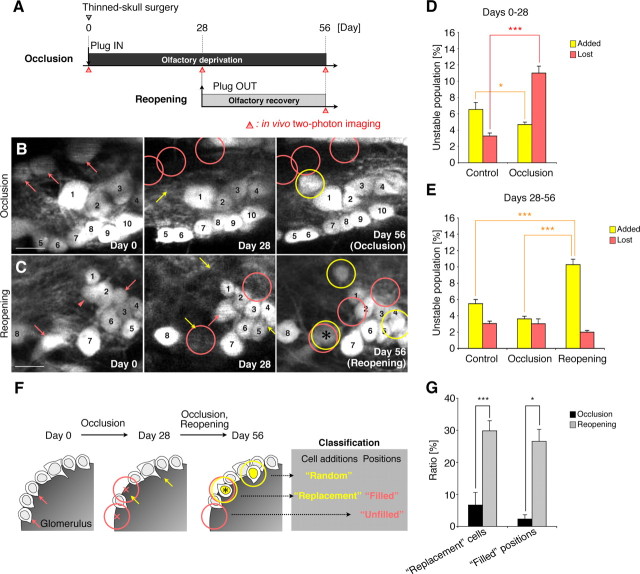
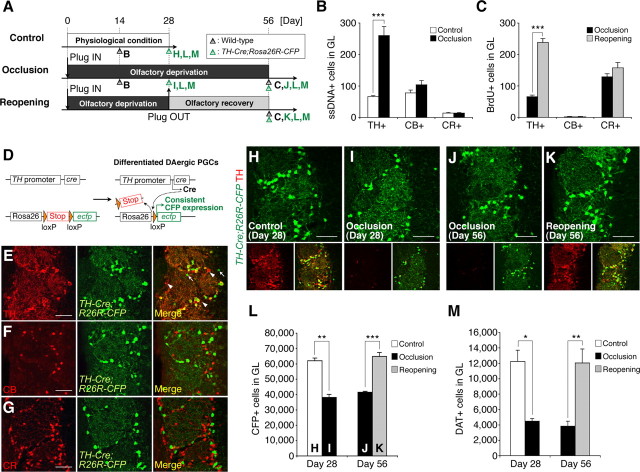
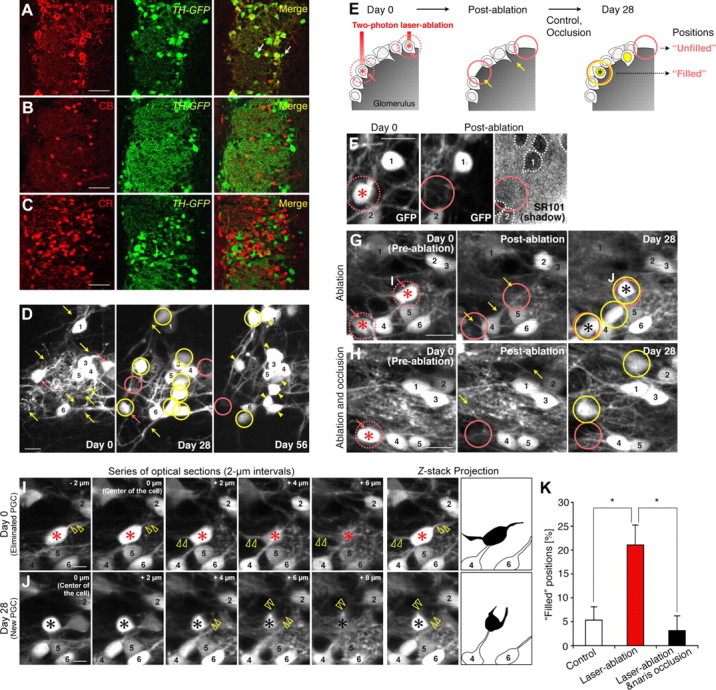
Throughout life, new neurons are added and old ones eliminated in the adult mouse olfactory bulb. Previous studies suggested that olfactory experience controls the process by which new neurons are integrated into mature circuits. Here we report novel olfactory-experience-dependent mechanisms of neuronal turnover. Using two-photon laser-scanning microscopy and sensory manipulations in adult live mice, we found that the neuronal turnover was dynamically controlled by olfactory input in a neuronal subtype-specific manner. Olfactory input enhanced this turnover, which was characterized by the reiterated use of the same positions in the glomeruli by new neurons. Our results suggest that olfactory-experience-dependent modification of neuronal turnover confers structural plasticity and stability on the olfactory bulb.
Figures
Similar articles
-
A subtype-specific critical period for neurogenesis in the postnatal development of mouse olfactory glomeruli.PLoS One. 2012;7(11):e48431. doi: 10.1371/journal.pone.0048431. Epub 2012 Nov 1. PLoS One. 2012. PMID: 23133633 Free PMC article.
-
Dense representation of natural odorants in the mouse olfactory bulb.Nat Neurosci. 2012 Mar 11;15(4):537-9. doi: 10.1038/nn.3057. Nat Neurosci. 2012. PMID: 22406552 Free PMC article.
-
Turnover of newborn olfactory bulb neurons optimizes olfaction.J Neurosci. 2009 Sep 30;29(39):12302-14. doi: 10.1523/JNEUROSCI.3383-09.2009. J Neurosci. 2009. PMID: 19793989 Free PMC article.
-
Newborn neurons in the adult olfactory bulb: unique properties for specific odor behavior.Behav Brain Res. 2012 Feb 14;227(2):480-9. doi: 10.1016/j.bbr.2011.08.001. Epub 2011 Aug 6. Behav Brain Res. 2012. PMID: 21843557 Review.
-
The olfactory bulb: coding and processing of odor molecule information.Science. 1999 Oct 22;286(5440):711-5. doi: 10.1126/science.286.5440.711. Science. 1999. PMID: 10531048 Review.
Cited by
-
Excitable Axonal Domains Adapt to Sensory Deprivation in the Olfactory System.J Neurosci. 2022 Feb 23;42(8):1491-1509. doi: 10.1523/JNEUROSCI.0305-21.2021. Epub 2022 Jan 12. J Neurosci. 2022. PMID: 35022219 Free PMC article.
-
Functional Maturation and Experience-Dependent Plasticity in Adult-Born Olfactory Bulb Dopaminergic Neurons.Eur J Neurosci. 2025 Jul;62(1):e70188. doi: 10.1111/ejn.70188. Eur J Neurosci. 2025. PMID: 40653358 Free PMC article.
-
Cell type- and layer-specific plasticity of olfactory bulb interneurons following olfactory sensory neuron ablation.Sci Rep. 2024 Aug 1;14(1):17771. doi: 10.1038/s41598-024-68649-4. Sci Rep. 2024. PMID: 39090136 Free PMC article.
-
Adult Born Olfactory Bulb Dopaminergic Interneurons: Molecular Determinants and Experience-Dependent Plasticity.Front Neurosci. 2016 May 6;10:189. doi: 10.3389/fnins.2016.00189. eCollection 2016. Front Neurosci. 2016. PMID: 27199651 Free PMC article. Review.
-
The Functional Role of Olfactory Bulb Granule Cell Subtypes Derived From Embryonic and Postnatal Neurogenesis.Front Mol Neurosci. 2018 Jul 5;11:229. doi: 10.3389/fnmol.2018.00229. eCollection 2018. Front Mol Neurosci. 2018. PMID: 30034321 Free PMC article. Review.
References
-
- Altman J. Autoradiographic and histological studies of postnatal neurogenesis. IV. Cell proliferation and migration in the anterior forebrain, with special reference to persisting neurogenesis in the olfactory bulb. J Comp Neurol. 1969;137:433–457. - PubMed
-
- Alvarez-Buylla A, Lim DA. For the long run: maintaining germinal niches in the adult brain. Neuron. 2004;41:683–686. - PubMed
-
- Baker H, Morel K, Stone DM, Maruniak JA. Adult naris closure profoundly reduces tyrosine hydroxylase expression in mouse olfactory bulb. Brain Res. 1993;614:109–116. - PubMed
-
- Bastien-Dionne PO, David LS, Parent A, Saghatelyan A. Role of sensory activity on chemospecific populations of interneurons in the adult olfactory bulb. J Comp Neurol. 2010;518:1847–1861. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials