

Degeneracy and neuromodulation among thermosensory neurons contribute to robust thermosensory behaviors in Caenorhabditis elegans
- PMID: 21832201
- PMCID: PMC3167209
- DOI: 10.1523/JNEUROSCI.1098-11.2011
Degeneracy and neuromodulation among thermosensory neurons contribute to robust thermosensory behaviors in Caenorhabditis elegans
Abstract
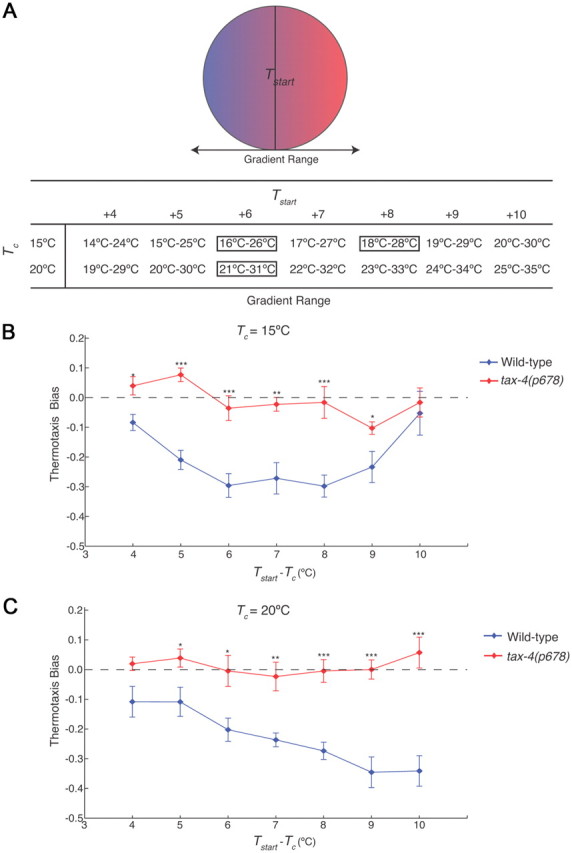
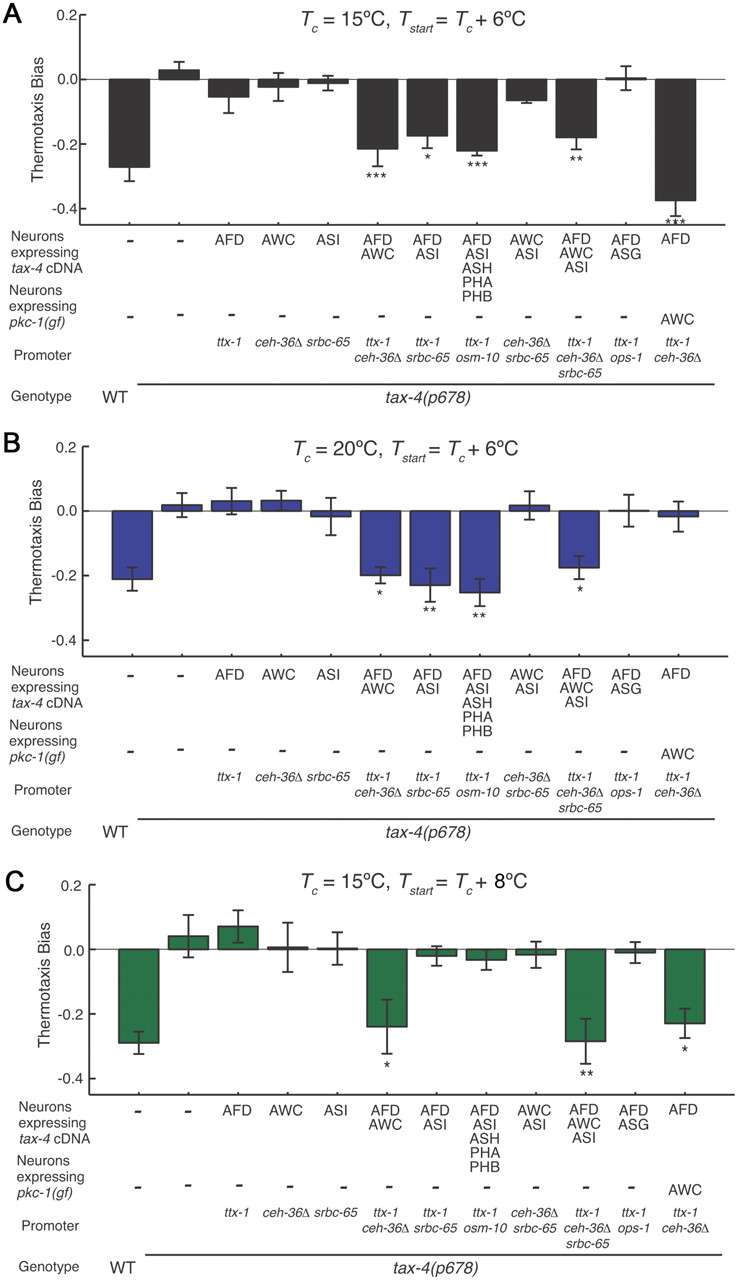
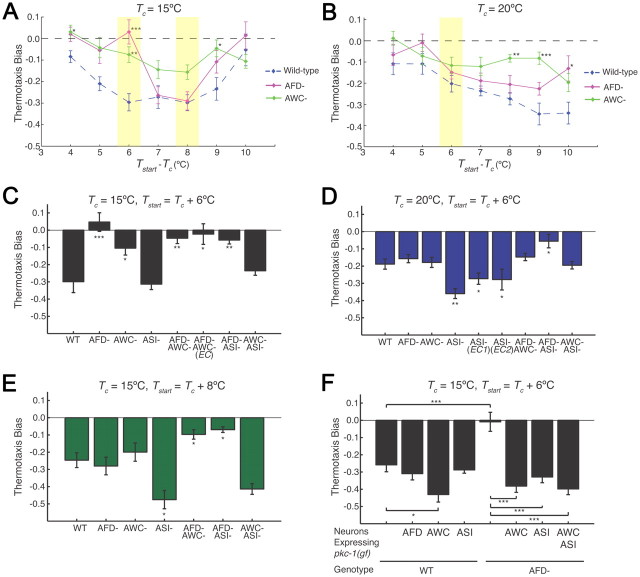
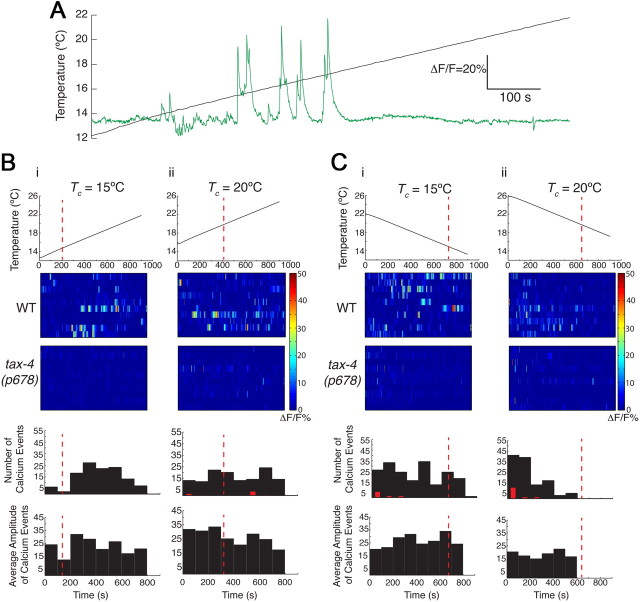
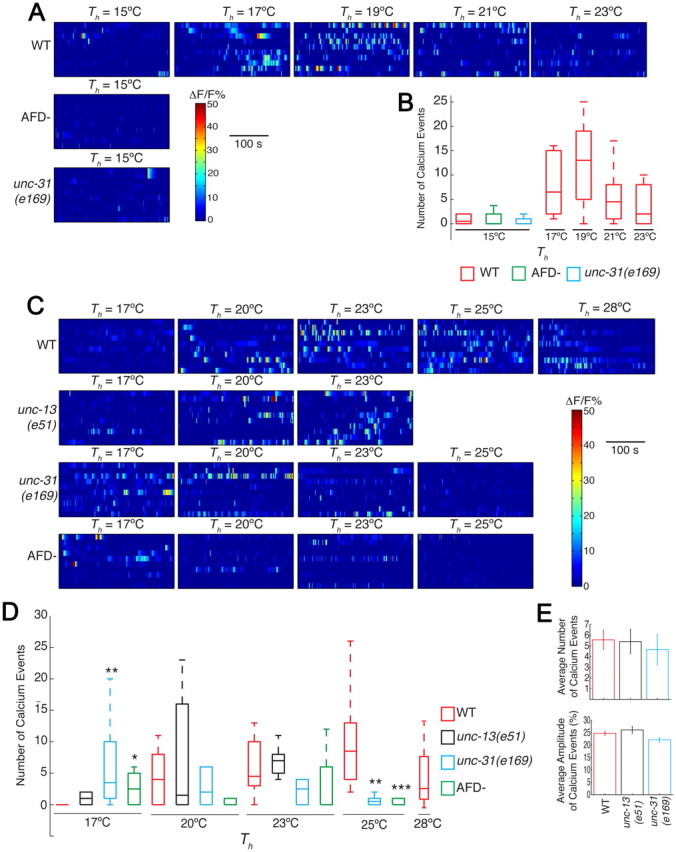
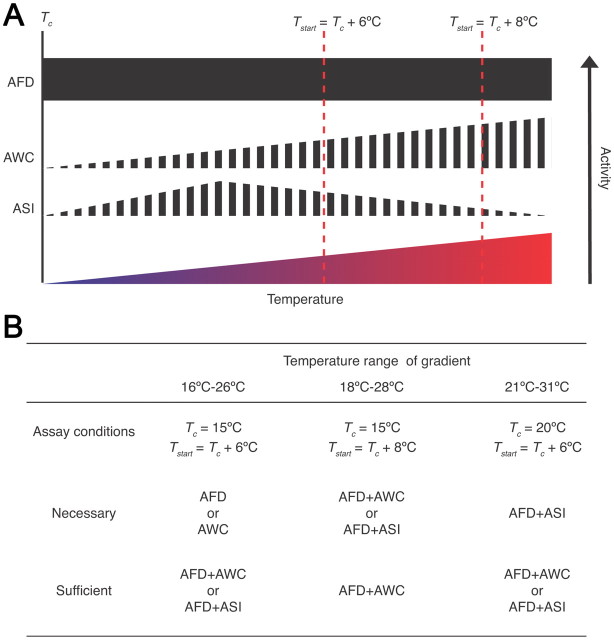
Animals must ensure that they can execute behaviors important for physiological homeostasis under constantly changing environmental conditions. The neural mechanisms that regulate this behavioral robustness are not well understood. The nematode Caenorhabditis elegans thermoregulates primarily via modulation of navigation behavior. Upon encountering temperatures higher than its cultivation temperature (T(c)), C. elegans exhibits negative thermotaxis toward colder temperatures using a biased random walk strategy. We find that C. elegans exhibits robust negative thermotaxis bias under conditions of varying T(c) and temperature ranges. By cell ablation and cell-specific rescue experiments, we show that the ASI chemosensory neurons are newly identified components of the thermosensory circuit, and that different combinations of ASI and the previously identified AFD and AWC thermosensory neurons are necessary and sufficient under different conditions to execute a negative thermotaxis strategy. ASI responds to temperature stimuli within a defined operating range defined by T(c), and signaling from AFD regulates the bounds of this operating range, suggesting that neuromodulation among thermosensory neurons maintains coherence of behavioral output. Our observations demonstrate that a negative thermotaxis navigational strategy can be generated via different combinations of thermosensory neurons acting degenerately, and emphasize the importance of defining context when analyzing neuronal contributions to a behavior.
Figures
Comment in
-
Structural basis for degeneracy among thermosensory neurons in Caenorhabditis elegans.J Neurosci. 2012 Jan 4;32(1):1-3. doi: 10.1523/JNEUROSCI.5112-11.2012. J Neurosci. 2012. PMID: 22219264 Free PMC article. No abstract available.
References
-
- Anderson JL, Albergotti L, Proulx S, Peden C, Huey RB, Phillips PC. Thermal preference of Caenorhabditis elegans: a null model and empirical tests. J Exp Biol. 2007;210:3107–3116. - PubMed
-
- Ann K, Kowalchyk JA, Loyet KM, Martin TF. Novel Ca2+-binding protein (CAPS) related to UNC-31 required for Ca2+-activated exocytosis. J Biol Chem. 1997;272:19637–19640. - PubMed
-
- Bargmann CI, Horvitz HR. Control of larval development by chemosensory neurons in Caenorhabditis elegans. Science. 1991;251:1243–1246. - PubMed
-
- Bargmann CI, Hartwieg E, Horvitz HR. Odorant-selective genes and neurons mediate olfaction in C. elegans. Cell. 1993;74:515–527. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources