

Tetraspanin CD151 maintains vascular stability by balancing the forces of cell adhesion and cytoskeletal tension
- PMID: 21832275
- PMCID: PMC3204742
- DOI: 10.1182/blood-2011-03-339531
Tetraspanin CD151 maintains vascular stability by balancing the forces of cell adhesion and cytoskeletal tension
Erratum in
- Blood. 2014 Jun 12;123(24):3843
Abstract
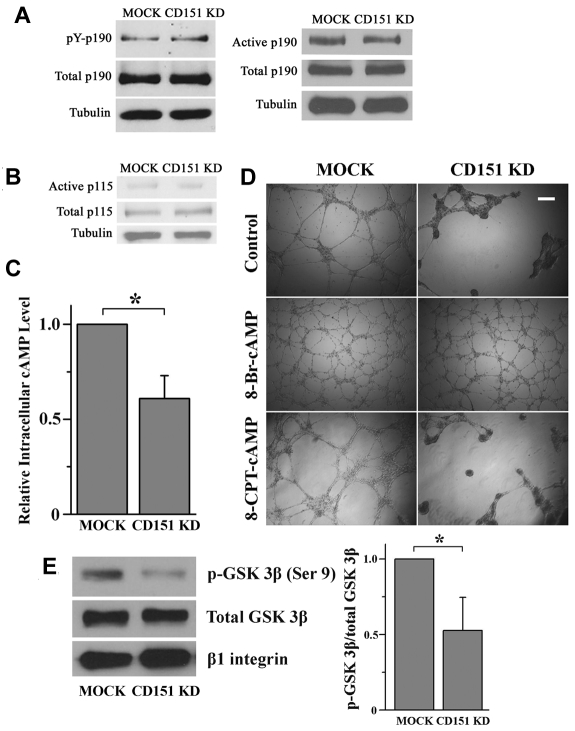
Tetraspanin CD151 is highly expressed in endothelial cells and regulates pathologic angiogenesis. However, the mechanism by which CD151 promotes vascular morphogenesis and whether CD151 engages other vascular functions are unclear. Here we report that CD151 is required for maintaining endothelial capillary-like structures formed in vitro and the integrity of endothelial cell-cell and cell-matrix contacts in vivo. In addition, vascular permeability is markedly enhanced in the absence of CD151. As a global regulator of endothelial cell-cell and cell-matrix adhesions, CD151 is needed for the optimal functions of various cell adhesion proteins. The loss of CD151 elevates actin cytoskeletal traction by up-regulating RhoA signaling and diminishes actin cortical meshwork by down-regulating Rac1 activity. The inhibition of RhoA or activation of cAMP signaling stabilizes CD151-silenced or -null endothelial structure in vascular morphogenesis. Together, our data demonstrate that CD151 maintains vascular stability by promoting endothelial cell adhesions, especially cell-cell adhesion, and confining cytoskeletal tension.
Figures





References
-
- Sterk LM, Geuijen CA, van den Berg JG, Claessen N, Weening JJ, Sonnenberg A. Association of the tetraspanin CD151 with the laminin-binding integrins alpha3beta1, alpha6beta1, alpha6beta4 and alpha7beta1 in cells in culture and in vivo. J Cell Sci. 2002;115(6):1161–1173. - PubMed
-
- Sincock PM, Fitter S, Parton RG, Berndt MC, Gamble JR, Ashman LK. PETA-3/CD151, a member of the transmembrane 4 superfamily, is localised to the plasma membrane and endocytic system of endothelial cells, associates with multiple integrins and modulates cell function. J Cell Sci. 1999;112(6):833–844. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
