

Evidence from the agonadal juvenile male rhesus monkey (Macaca mulatta) for the view that the action of neurokinin B to trigger gonadotropin-releasing hormone release is upstream from the kisspeptin receptor
- PMID: 21832818
- PMCID: PMC3238032
- DOI: 10.1159/000329045
Evidence from the agonadal juvenile male rhesus monkey (Macaca mulatta) for the view that the action of neurokinin B to trigger gonadotropin-releasing hormone release is upstream from the kisspeptin receptor
Abstract
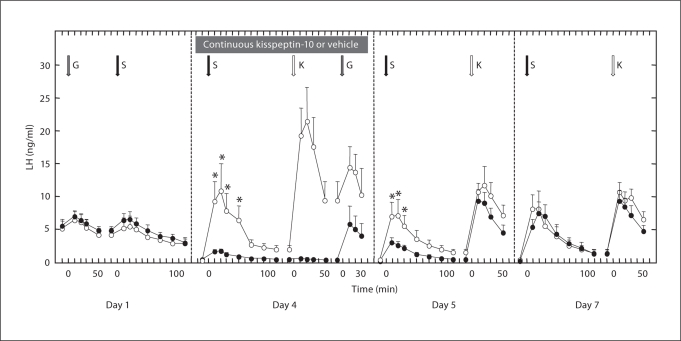
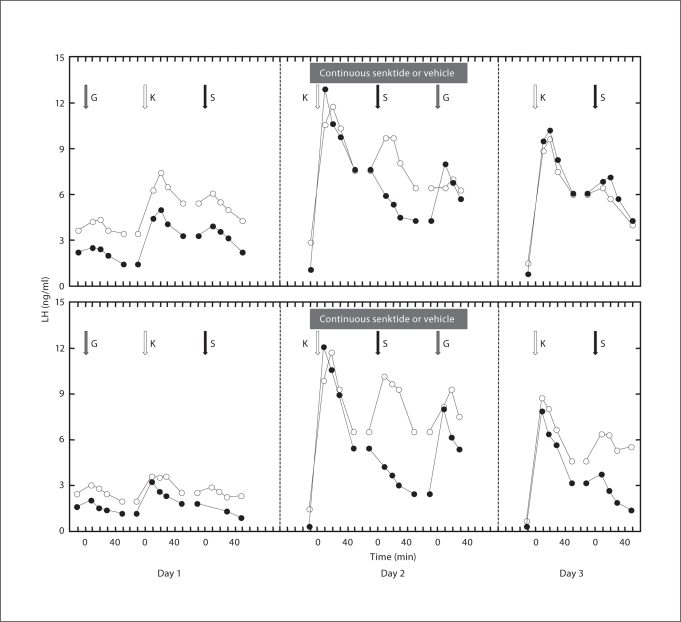
Human genetics have revealed that kisspeptin signaling and neurokinin B (NKB) signaling are both required for robust pulsatile gonadotropin-releasing hormone (GnRH) release, and therefore for puberty and maintenance of adult gonadal function. How these two peptides interact to affect GnRH pulse generation remains a mystery. To address the hierarchy of the NKB and kisspeptin signaling pathways that are essential for GnRH release, two experiments were conducted using agonadal, juvenile male monkeys. Pituitary responsiveness to GnRH was first heightened by a pulsatile GnRH infusion to use the in situ pituitary as a bioassay for GnRH release. In the first experiment (n = 3), the kisspeptin receptor (KISS1R) was desensitized by a continuous 99-hour i.v. infusion of kisspeptin-10 (100 μg/h). During the last 4 h of continuous kisspeptin-10 infusion, desensitization of KISS1R was confirmed by failure of an i.v. bolus of kisspeptin-10 to elicit GnRH release. Desensitization of KISS1R was associated with a markedly blunted GnRH response to senktide. The response to senktide was progressively restored during the 72 h following termination of continuous kisspeptin-10. An analogous design was employed in the second experiment (n = 2) to desensitize the NKB receptor (neurokinin 3 receptor, NK3R) by administration of a continuous 48-hour i.v. infusion of senktide (200 μg/h). While a bolus of senktide during the last 3 h of continuous senktide administration failed to elicit GnRH release, thus confirming desensitization of NK3R, the ability of kisspeptin to stimulate GnRH was unimpaired. The foregoing findings support the view that NKB stimulation of GnRH release is upstream from KISS1R.
Copyright © 2011 S. Karger AG, Basel.
Figures
References
-
- Seminara SB, Messager S, Chatzidaki EE, Thresher RR, Acierno JS, Jr, Shagoury JK, Bo-Abbas Y, Kuohung W, Schwinof KM, Hendrick A, Zahn D, Dixon J, Kaiser UB, Slaugenhaupt SA, Gusella JF, O'Rahilly S, Carlton MB, Crowley W, Jr, Apaicio SA, Colledge WH. The GPR54 gene as a regulator of puberty. N Engl J Med. 2003;349:1614–1627. - PubMed
-
- Topaloglu AK, Reimann F, Guclu M, Yalin AS, Kotan LD, Porter KM, Serin A, Mungan NL, Cook JR, Ozbek MN, Imamoglu S, Akalin NS, Yuksel B, O'Rahilly S, Semple RK. TAC3 and TACR3 mutations in familial hypogonadotropic hypogonadism reveal a key role for neurokinin B in the central control of reproduction. Nat Genet. 2009;41:354–358. - PMC - PubMed
-
- Guran T, Tolhurst G, Bereket A, Rocha N, Porter K, Turan S, Gribble FM, Kotan LD, Akcay T, Atay Z, Canan H, Serin A, O'Rahilly S, Reimann F, Semple RK, Topaloglu AK. Hypogonadotropic hypogonadism due to a novel missense mutation in the first extracellular loop of the neurokinin B receptor. J Clin Endocrinol Metab. 2009;94:3633–3639. - PMC - PubMed
-
- Young J, Bouligand J, Francou B, Raffin-Sanson ML, Gaillez S, Jeanpierre M, Grynberg M, Kamenicky P, Chanson P, Brailly-Tabard S, Guiochon-Mantel A. TAC3 and TACR3 defects cause hypothalamic congenital hypogonadotropic hypogonadism in humans. J Clin Endocrinol Metab. 2010;95:2287–2295. - PubMed
