

Primary infection by a human immunodeficiency virus with atypical coreceptor tropism
- PMID: 21835785
- PMCID: PMC3187499
- DOI: 10.1128/JVI.05249-11
Primary infection by a human immunodeficiency virus with atypical coreceptor tropism
Abstract
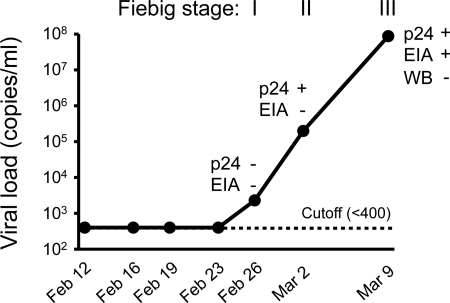
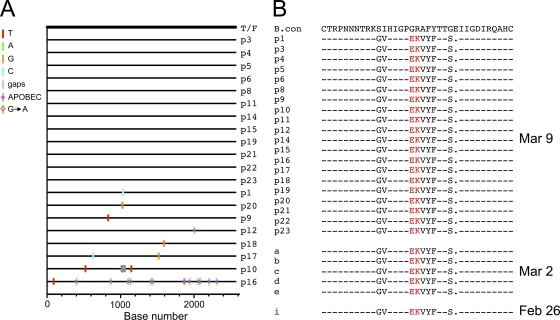
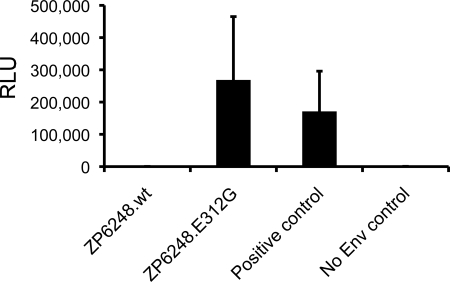
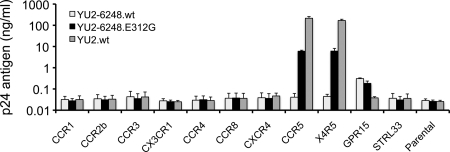
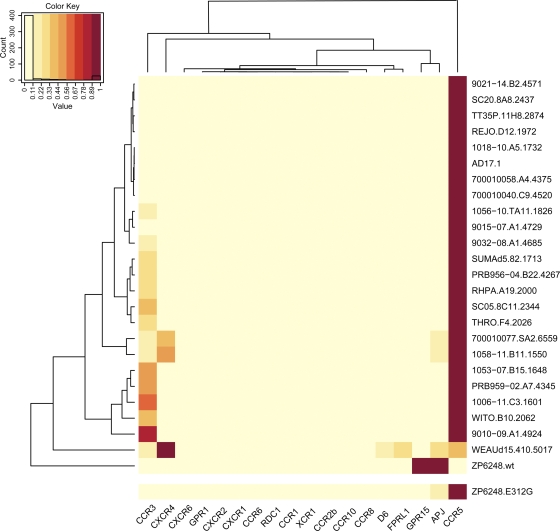
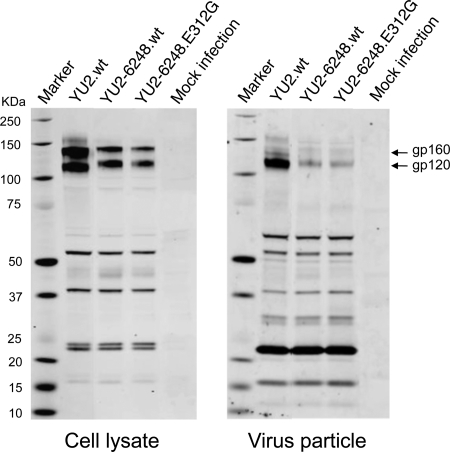
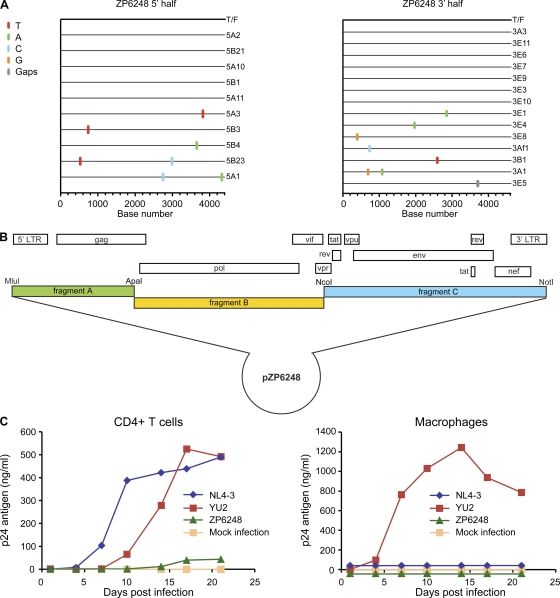
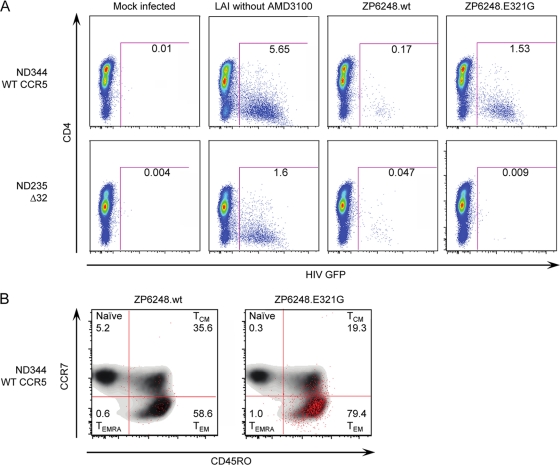
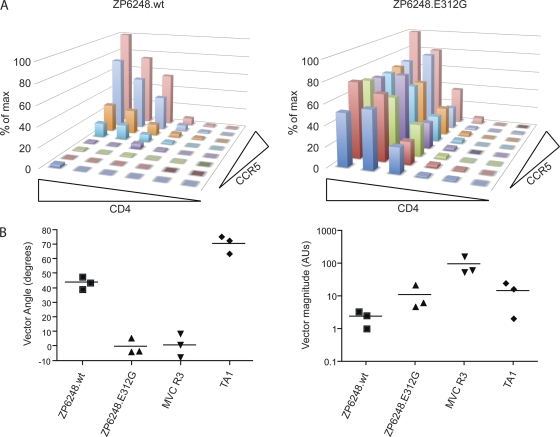
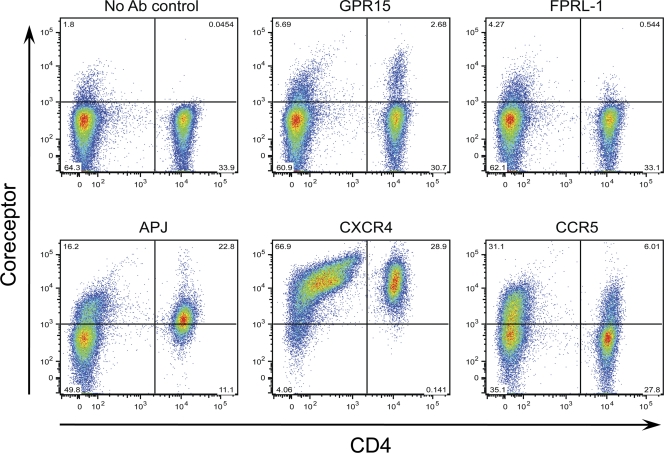
The great majority of human immunodeficiency virus type 1 (HIV-1) strains enter CD4+ target cells by interacting with one of two coreceptors, CCR5 or CXCR4. Here we describe a transmitted/founder (T/F) virus (ZP6248) that was profoundly impaired in its ability to utilize CCR5 and CXCR4 coreceptors on multiple CD4+ cell lines as well as primary human CD4+ T cells and macrophages in vitro yet replicated to very high titers (>80 million RNA copies/ml) in an acutely infected individual. Interestingly, the envelope (Env) glycoprotein of this clade B virus had a rare GPEK sequence in the crown of its third variable loop (V3) rather than the consensus GPGR sequence. Extensive sequencing of sequential plasma samples showed that the GPEK sequence was present in virtually all Envs, including those from the earliest time points after infection. The molecularly cloned (single) T/F virus was able to replicate, albeit poorly, in cells obtained from ccr5Δ32 homozygous donors. The ZP6248 T/F virus could also infect cell lines overexpressing the alternative coreceptors GPR15, APJ, and FPRL-1. A single mutation in the V3 crown sequence (GPEK->GPGK) of ZP6248 restored its infectivity in CCR5+ cells but reduced its ability to replicate in GPR15+ cells, indicating that the V3 crown motif played an important role in usage of this alternative coreceptor. These results suggest that the ZP6248 T/F virus established an acute in vivo infection by using coreceptor(s) other than CCR5 or CXCR4 or that the CCR5 coreceptor existed in an unusual conformation in this individual.
Figures
Similar articles
-
Linkages between HIV-1 specificity for CCR5 or CXCR4 and in vitro usage of alternative coreceptors during progressive HIV-1 subtype C infection.Retrovirology. 2013 Sep 16;10:98. doi: 10.1186/1742-4690-10-98. Retrovirology. 2013. PMID: 24041034 Free PMC article.
-
The variable loop 3 in the envelope glycoprotein is critical for the atypical coreceptor usage of an HIV-1 strain.PLoS One. 2014 Jun 4;9(6):e98058. doi: 10.1371/journal.pone.0098058. eCollection 2014. PLoS One. 2014. PMID: 24897520 Free PMC article.
-
Use of GPR1, GPR15, and STRL33 as coreceptors by diverse human immunodeficiency virus type 1 and simian immunodeficiency virus envelope proteins.Virology. 1998 Sep 30;249(2):367-78. doi: 10.1006/viro.1998.9306. Virology. 1998. PMID: 9791028
-
HIV-1 coreceptor preference is distinct from target cell tropism: a dual-parameter nomenclature to define viral phenotypes.J Leukoc Biol. 2006 Nov;80(5):965-72. doi: 10.1189/jlb.0306148. Epub 2006 Aug 21. J Leukoc Biol. 2006. PMID: 16923919 Review.
-
The role of viral coreceptors and enhanced macrophage tropism in human immunodeficiency virus type 1 disease progression.Sex Health. 2004;1(1):23-34. doi: 10.1071/sh03006. Sex Health. 2004. PMID: 16335478 Review.
Cited by
-
CD300lf is the primary physiologic receptor of murine norovirus but not human norovirus.PLoS Pathog. 2020 Apr 6;16(4):e1008242. doi: 10.1371/journal.ppat.1008242. eCollection 2020 Apr. PLoS Pathog. 2020. PMID: 32251490 Free PMC article.
-
Viremic control and viral coreceptor usage in two HIV-1-infected persons homozygous for CCR5 Δ32.AIDS. 2015 May 15;29(8):867-76. doi: 10.1097/QAD.0000000000000629. AIDS. 2015. PMID: 25730507 Free PMC article.
-
HIV Associated Neurodegenerative Disorders: A New Perspective on the Role of Lipid Rafts in Gp120-Mediated Neurotoxicity.Curr HIV Res. 2018;16(4):258-269. doi: 10.2174/1570162X16666181003144740. Curr HIV Res. 2018. PMID: 30280668 Free PMC article. Review.
-
Development of a contemporary globally diverse HIV viral panel by the EQAPOL program.J Immunol Methods. 2014 Jul;409:117-30. doi: 10.1016/j.jim.2014.01.004. Epub 2014 Jan 19. J Immunol Methods. 2014. PMID: 24447533 Free PMC article.
-
HIV-2 interaction with cell coreceptors: amino acids within the V1/V2 region of viral envelope are determinant for CCR8, CCR5 and CXCR4 usage.Retrovirology. 2014 Nov 25;11:99. doi: 10.1186/s12977-014-0099-3. Retrovirology. 2014. PMID: 25421818 Free PMC article.
References
-
- Begaud E., et al. 2003. Broad spectrum of coreceptor usage and rapid disease progression in HIV-1-infected individuals from Central African Republic. AIDS Res. Hum. Retroviruses 19:551–560 - PubMed
-
- Berger E. A., Murphy P. M., Farber J. M. 1999. Chemokine receptors as HIV-1 coreceptors: roles in viral entry, tropism, and disease. Annu. Rev. Immunol. 17:657–700 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
