

Duplex unwinding and ATPase activities of the DEAD-box helicase eIF4A are coupled by eIF4G and eIF4B
- PMID: 21840318
- PMCID: PMC3175293
- DOI: 10.1016/j.jmb.2011.08.004
Duplex unwinding and ATPase activities of the DEAD-box helicase eIF4A are coupled by eIF4G and eIF4B
Abstract
Eukaryotic initiation factor (eIF) 4A is a DEAD-box helicase that stimulates translation initiation by unwinding mRNA secondary structure. The accessory proteins eIF4G, eIF4B, and eIF4H enhance the duplex unwinding activity of eIF4A, but the extent to which they modulate eIF4A activity is poorly understood. Here, we use real-time fluorescence assays to determine the kinetic parameters of duplex unwinding and ATP hydrolysis by these initiation factors. To ensure efficient duplex unwinding, eIF4B and eIF4G cooperatively activate the duplex unwinding activity of eIF4A. Our data reveal that eIF4H is much less efficient at stimulating eIF4A unwinding activity than eIF4B, implying that eIF4H is not able to completely substitute for eIF4B in duplex unwinding. By monitoring unwinding and ATPase assays under identical conditions, we demonstrate that eIF4B couples the ATP hydrolysis cycle of eIF4A with strand separation, thereby minimizing nonproductive unwinding events. Using duplex substrates with altered GC contents but similar predicted thermal stabilities, we further show that the rate of formation of productive unwinding complexes is strongly influenced by the local stability per base pair, in addition to the stability of the entire duplex. This finding explains how a change in the GC content of a hairpin is able to influence translation initiation while maintaining the overall predicted thermal stability.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
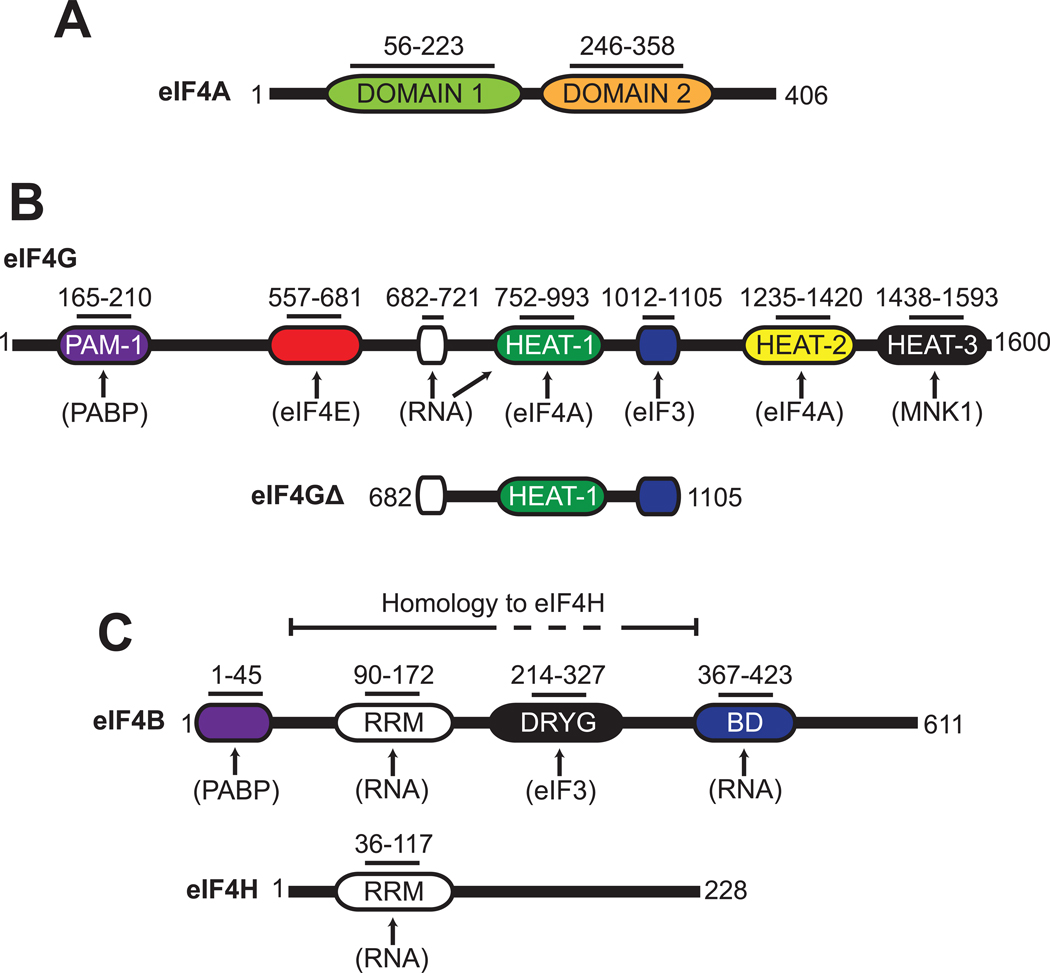
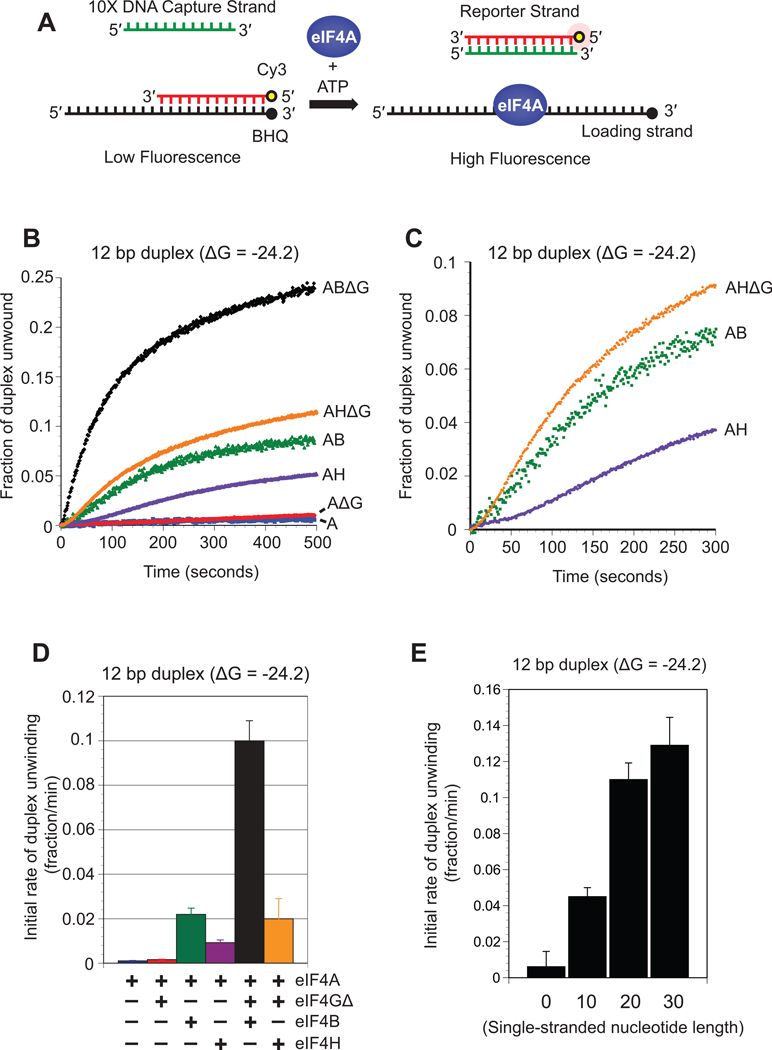
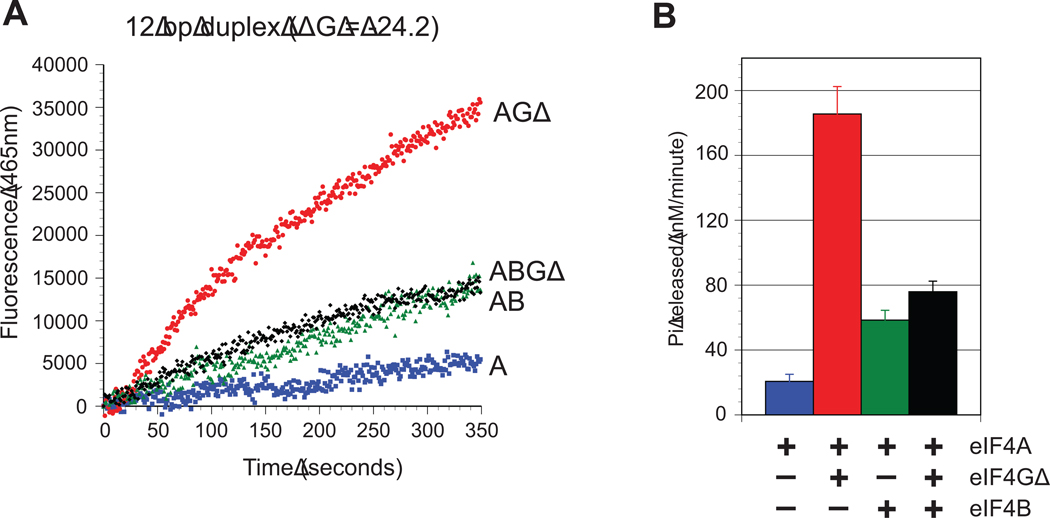
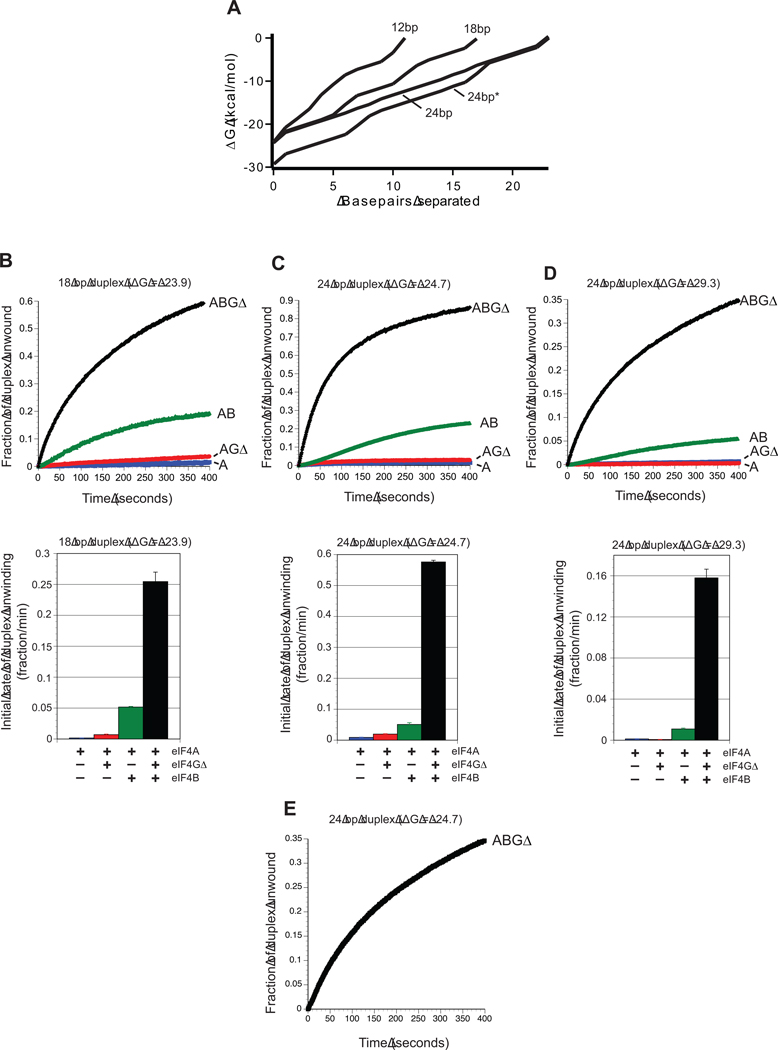
References
-
- Rogers GW, Jr, Richter NJ, Merrick WC. Biochemical and kinetic characterization of the RNA helicase activity of eukaryotic initiation factor 4A. J Biol Chem. 1999;274:12236–12244. - PubMed
-
- Rogers GW, Jr, Lima WF, Merrick WC. Further characterization of the helicase activity of eIF4A. Substrate specificity. J Biol Chem. 2001;276:12598–12608. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
