

Computational advances towards linking BOLD and behavior
- PMID: 21840553
- PMCID: PMC3384549
- DOI: 10.1016/j.neuropsychologia.2011.07.013
Computational advances towards linking BOLD and behavior
Abstract
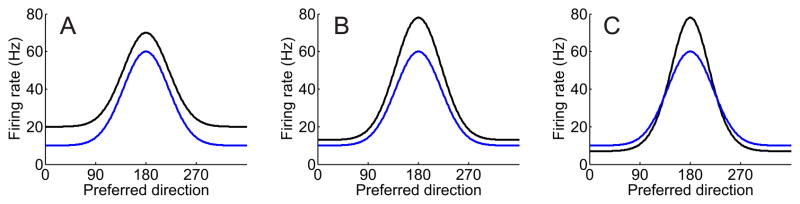
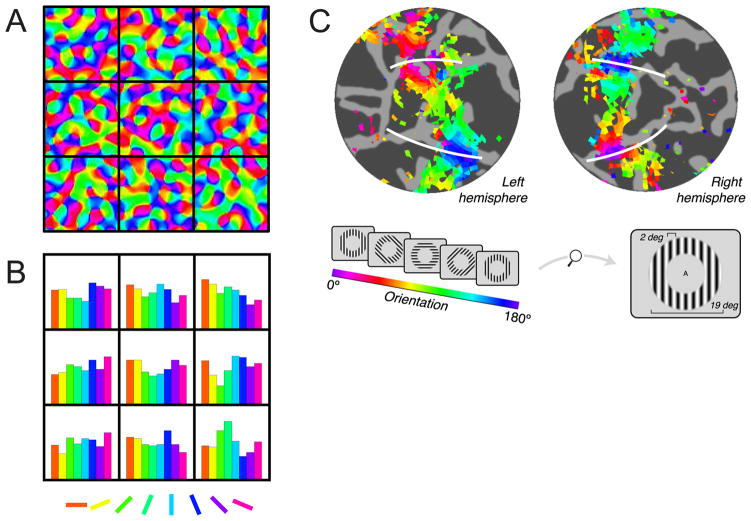
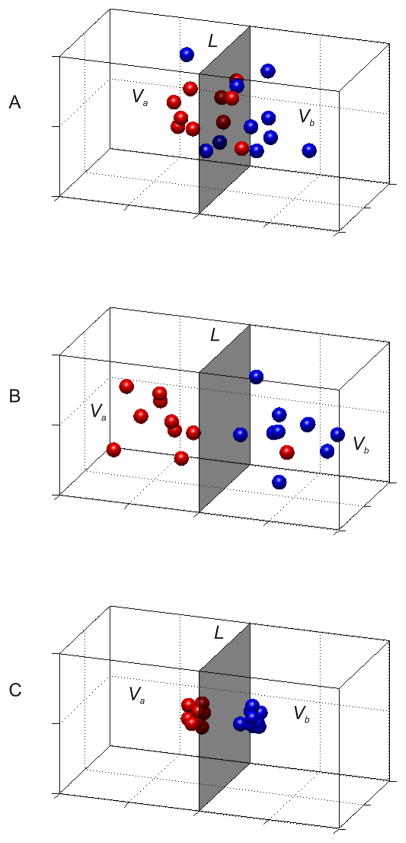
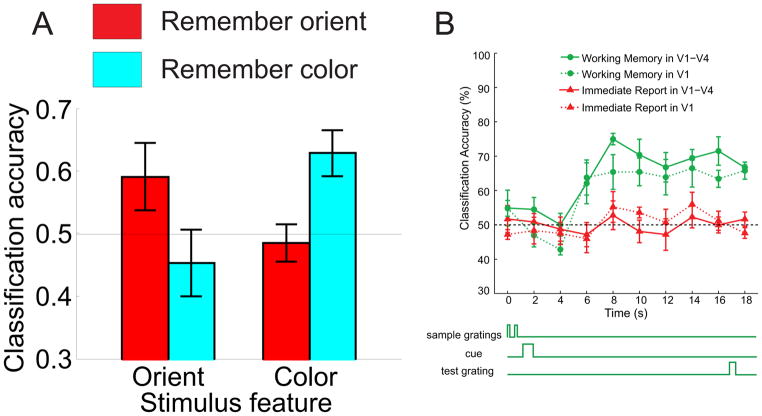
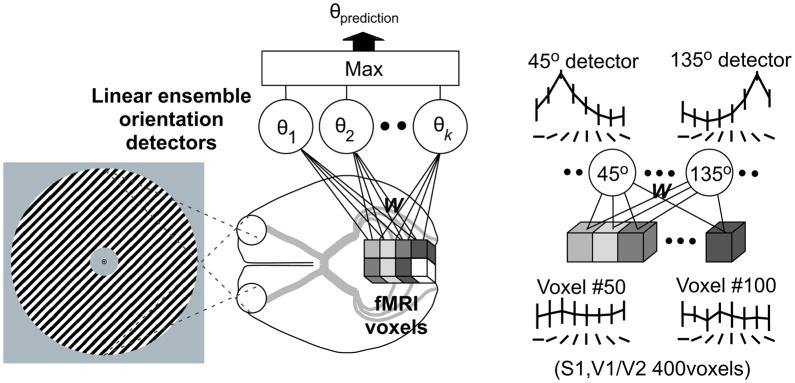
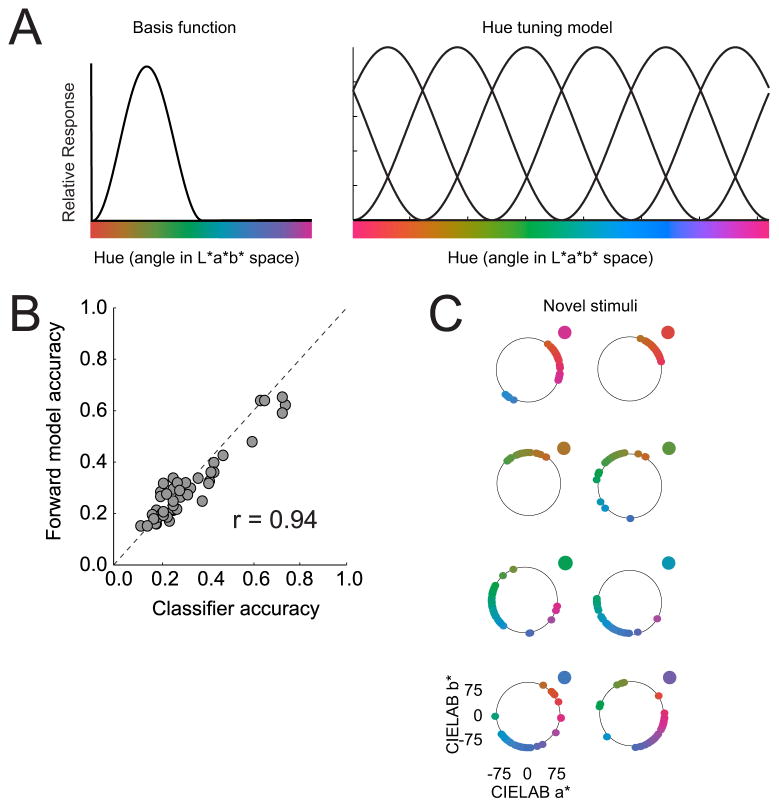
Traditionally, fMRI studies have focused on analyzing the mean response amplitude within a cortical area. However, the mean response is blind to many important patterns of cortical modulation, which severely limits the formulation and evaluation of linking hypotheses between neural activity, BOLD responses, and behavior. More recently, multivariate pattern classification analysis (MVPA) has been applied to fMRI data to evaluate the information content of spatially distributed activation patterns. This approach has been remarkably successful at detecting the presence of specific information in targeted brain regions, and provides an extremely flexible means of extracting that information without a precise generative model for the underlying neural activity. However, this flexibility comes at a cost: since MVPA relies on pooling information across voxels that are selective for many different stimulus attributes, it is difficult to infer how specific sub-sets of tuned neurons are modulated by an experimental manipulation. In contrast, recently developed encoding models can produce more precise estimates of feature-selective tuning functions, and can support the creation of explicit linking hypotheses between neural activity and behavior. Although these encoding models depend on strong - and often untested - assumptions about the response properties of underlying neural generators, they also provide a unique opportunity to evaluate population-level computational theories of perception and cognition that have previously been difficult to assess using either single-unit recording or conventional neuroimaging techniques.
Copyright © 2011. Published by Elsevier Ltd.
Figures
References
-
- Awh E, Jonides J. Overlapping mechanisms of attention and spatial working memory. Trends in cognitive sciences. 2001;5:119–126. - PubMed
-
- Boynton GM. Attention and visual perception. Current opinion in neurobiology. 2005a;15:465–469. - PubMed
-
- Boynton GM. Imaging orientation selectivity: decoding conscious perception in V1. Nature neuroscience. 2005b;8:541–542. - PubMed
-
- Boynton GM, Demb JB, Glover GH, Heeger DJ. Neuronal basis of contrast discrimination. Vision research. 1999;39:257–269. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
