

Actin dynamics counteract membrane tension during clathrin-mediated endocytosis
- PMID: 21841790
- PMCID: PMC3167020
- DOI: 10.1038/ncb2307
Actin dynamics counteract membrane tension during clathrin-mediated endocytosis
Abstract
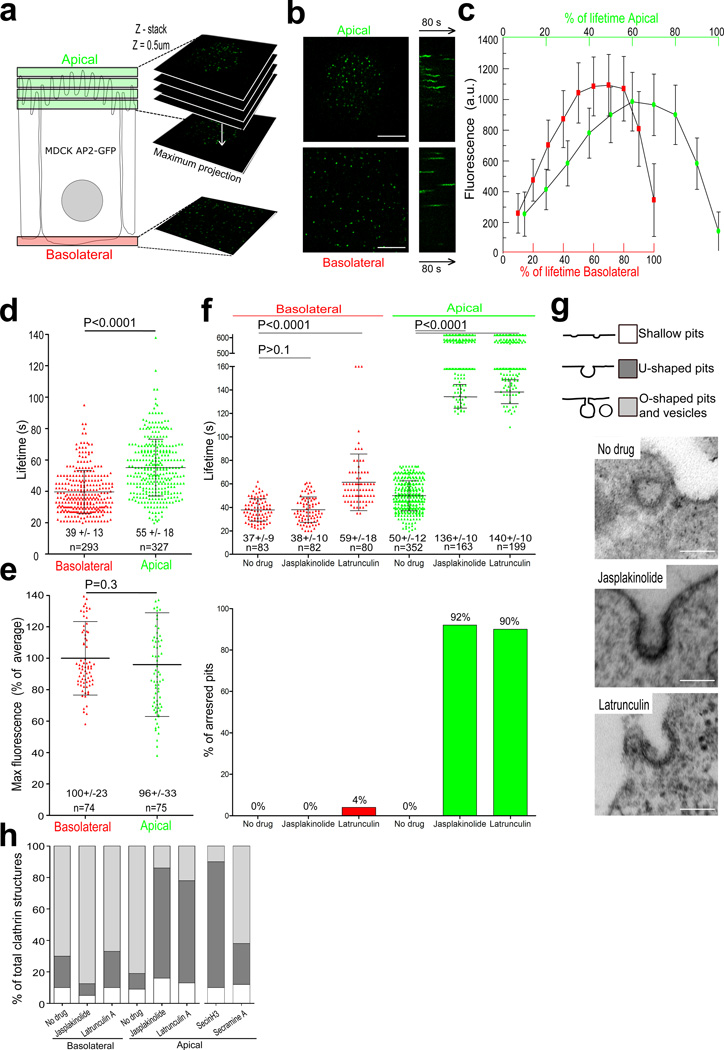
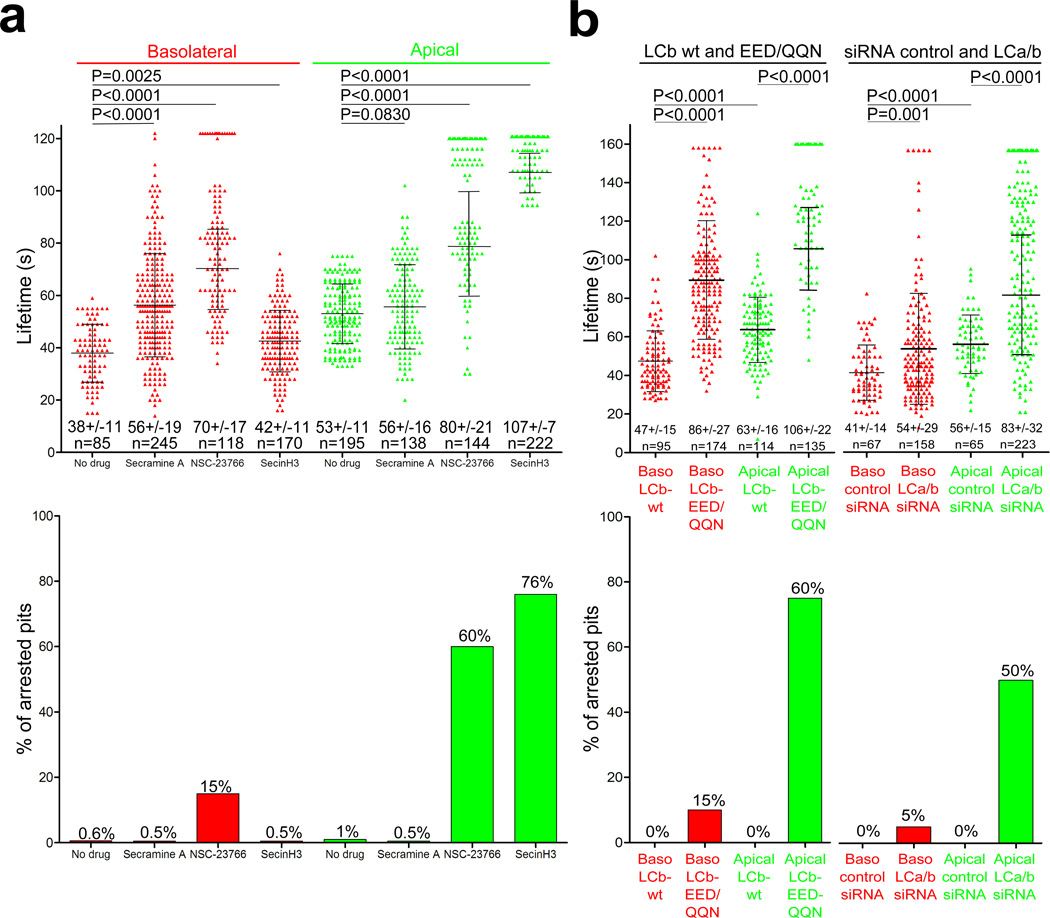
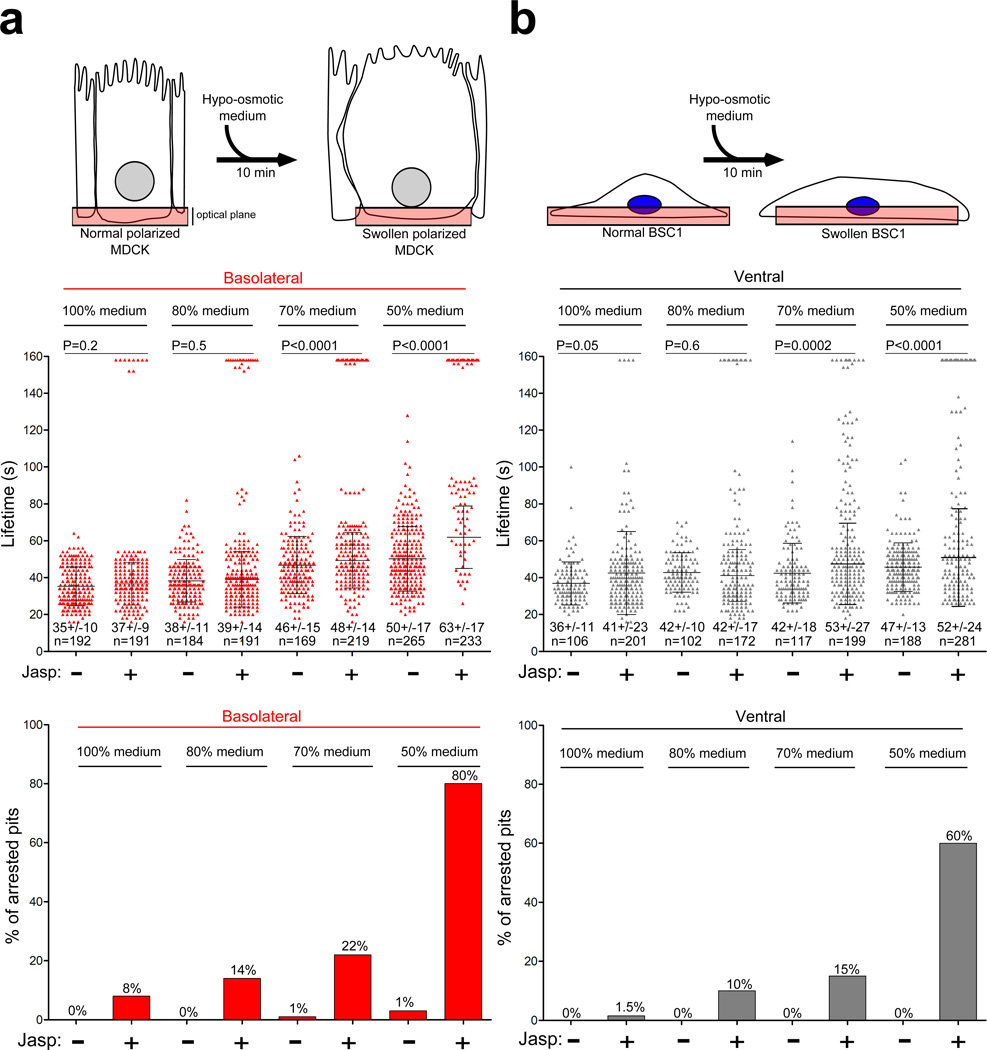
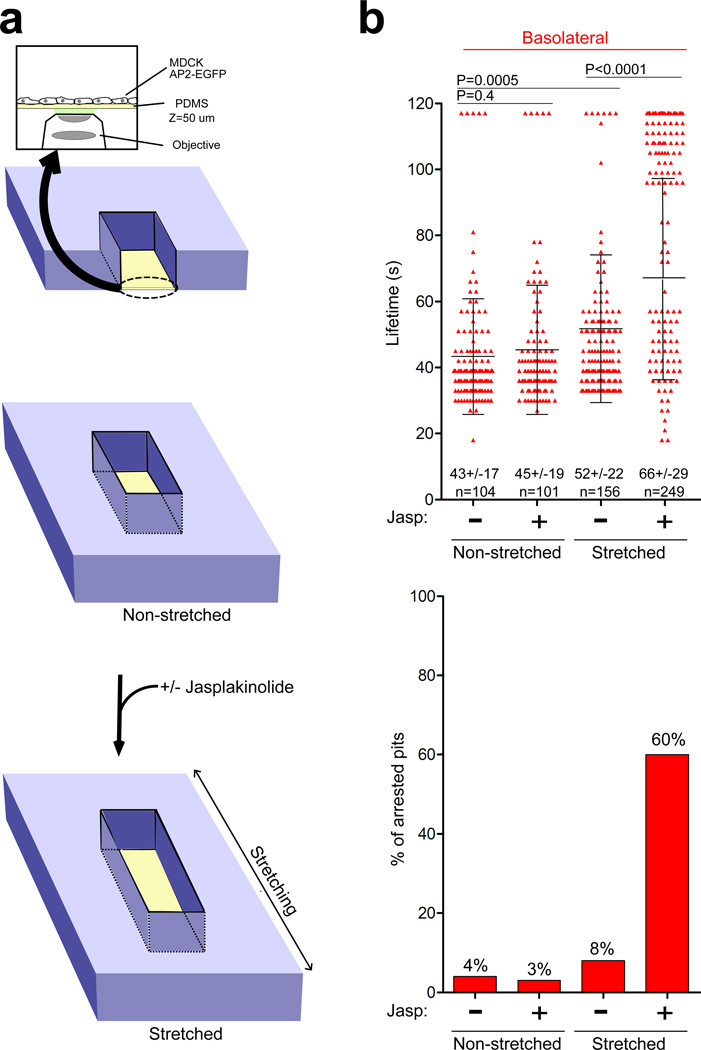
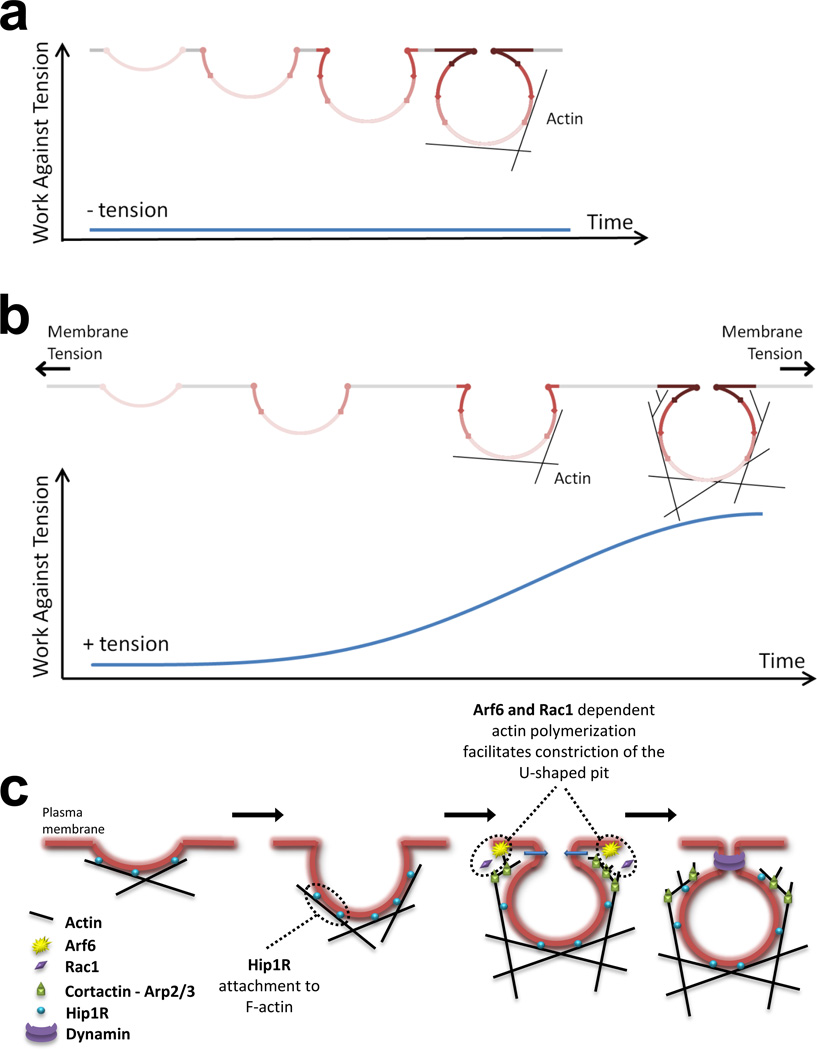
Clathrin-mediated endocytosis is independent of actin dynamics in many circumstances but requires actin polymerization in others. We show that membrane tension determines the actin dependence of clathrin-coat assembly. As found previously, clathrin assembly supports formation of mature coated pits in the absence of actin polymerization on both dorsal and ventral surfaces of non-polarized mammalian cells, and also on basolateral surfaces of polarized cells. Actin engagement is necessary, however, to complete membrane deformation into a coated pit on apical surfaces of polarized cells and, more generally, on the surface of any cell in which the plasma membrane is under tension from osmotic swelling or mechanical stretching. We use these observations to alter actin dependence experimentally and show that resistance of the membrane to propagation of the clathrin lattice determines the distinction between 'actin dependent and 'actin independent'. We also find that light-chain-bound Hip1R mediates actin engagement. These data thus provide a unifying explanation for the role of actin dynamics in coated-pit budding.
Figures
References
-
- Ehrlich M, et al. Endocytosis by random initiation and stabilization of clathrin-coated pits. Cell. 2004;118:591–605. - PubMed
-
- Conibear E. Converging views of endocytosis in yeast and mammals. Curr Opin Cell Biol. 2010;22:513–518. - PubMed
-
- Kaksonen M, Toret CP, Drubin DG. A modular design for the clathrin- and actin-mediated endocytosis machinery. Cell. 2005;123:305–320. - PubMed
-
- Merrifield CJ, Feldman ME, Wan L, Almers W. Imaging actin and dynamin recruitment during invagination of single clathrin-coated pits. Nature cell biology. 2002;4:691–698. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
